


- Home
- Volume 17 (2013)
- numéro 1
- Efficacité des méthodes de lutte contre le développement de cinq espèces de plantes invasives amphibies : Crassula helmsii, Hydrocotyle ranunculoides, Ludwigia grandiflora, Ludwigia peploides et Myriophyllum aquaticum (synthèse bibliographique)


View(s): 6414 (50 ULiège)
Download(s): 450 (8 ULiège)

Efficacité des méthodes de lutte contre le développement de cinq espèces de plantes invasives amphibies : Crassula helmsii, Hydrocotyle ranunculoides, Ludwigia grandiflora, Ludwigia peploides et Myriophyllum aquaticum (synthèse bibliographique)

Editor's Notes
Reçu le 3 février 2012, accepté le 8 novembre 2012
Résumé
Parmi les espèces invasives connaissant une expansion rapide en Europe, les plantes dites « amphibies » sont particulièrement problématiques. Elles sont capables de former des populations très denses à la surface des plans d’eau, se développant à la fois sous l’eau et hors de l’eau. La lutte contre ces espèces, en vue de l’éradication totale ou du ralentissement de l’invasion, représente un défi de taille pour les gestionnaires. Une synthèse bibliographique des méthodes de lutte existantes pour Crassula helmsii, Hydrocotyle ranunculoides, Ludwigia grandiflora, Ludwigia peploides et Myriophyllum aquaticum a été réalisée, prenant en considération leur efficacité et leur cout. Des 87 études analysées, il ressort une grande disparité d’effort de recherche (nombre d’études, échelles spatiales des études) entre méthodes de lutte et entre espèces. Néanmoins, l’efficacité des différentes méthodes de lutte est comparable pour les cinq espèces étudiées. Globalement, les cas d’éradication sont rares. Ils sont le résultat de luttes mécaniques ou chimiques, souvent couplées à des interventions complémentaires telles que la finition manuelle. Malgré une forte occurrence dans la littérature, la lutte chimique, moins fastidieuse à mettre en place et moins couteuse que les autres méthodes, n’a montré des niveaux d’efficacité satisfaisants qu’avec certaines matières actives. De même, la lutte biologique a montré peu de résultats encourageants à ce jour. Il est à noter que certains agents de lutte biologique sont d’ores et déjà considérés comme des espèces invasives en Europe. A contrario, malgré leur faible occurrence dans la littérature, l’arrachage manuel ou mécanique suivi de finitions manuelles a donné des niveaux d’efficacité satisfaisants. Vu les difficultés de lutter contre les espèces invasives amphibies et les dommages collatéraux potentiels sur les écosystèmes, un débat impliquant scientifiques, autorités et gestionnaires de plans d’eau devra avoir lieu afin de poser les bases d’une gestion cohérente à grande échelle.
Abstract
Effectiveness of techniques used to control the development of five invasive amphibious plant species: Crassula helmsii, Hydrocotyle ranunculoides, Ludwigia grandiflora, Ludwigia peploides and Myriophyllum aquaticum. A review. Among rapidly spreading invasive species in Europe, amphibious plants are particularly problematic. These plants are capable of developing a photosynthetic activity both below and above the water surface, and they typically form dense floating populations. Controlling amphibious plants, whether for the purposes of total eradication or for the regulation of their spread, represents an important challenge for managers. A literature review of existing control methods for Crassula helmsii, Hydrocotyle ranunculoides, Ludwigia grandiflora, Ludwigia peploides and Myriophyllum aquaticum was carried out, taking into account efficiency and costs. Of the 87 studies analyzed, a great variation was evident in the research efforts undertaken (number and spatial scale of studies) between species and between control methods. However, overall, the different control methods described showed a similar level of efficiency between species. Reports of total eradication of amphibious plants were rare. Such cases resulted from the application of mechanical or chemical methods, most often coupled with complementary methods such as manual finishing. The use of chemical control methods (less laborious and costly than others) was found to be frequently documented in the literature. However, except in the case of some active substances, such chemical methods were found to provide low control efficiency. Similarly, to date, biological control has not been found to provide convincing results. In addition, it should be noted that several biological control agents are considered as invasive species in Europe. On the other hand, although little related research has been carried out, methods such as manual or mechanical removal with manual finishing have often been found to result in satisfactory control levels. Aware of the difficulty of controlling amphibious plants, as well as the potential collateral damage caused by control methods to aquatic ecosystems, we argue that consensus still needs to be found between scientists, decision-makers and managers before decisions at regional or national scale can be made.
Table of content
1. Introduction
1Les espèces végétales aquatiques invasives des eaux douces connaissent une expansion grandissante en Europe. Parmi celles-ci, on distingue les formes de vie totalement immergées des espèces dites « amphibies », i.e. qui développent une activité photosynthétique à la fois sous l’eau et hors de l’eau. Ces dernières forment typiquement des populations denses à la surface des plans d’eaux, modifiant profondément le fonctionnement des écosystèmes aquatiques. Cinq espèces amphibies sont reprises dans la « liste noire des espèces invasives » du système d’information Harmonia, développé en Belgique sur base des impacts qu’ont les espèces invasives sur les écosystèmes (Branquart, 2009) et régulièrement prises en exemple dans le reste de l’Europe. Ces cinq mêmes espèces amphibies se retrouvent d’ailleurs dans la plupart des listes de plantes invasives des pays européens (Verbrugge et al., 2012). Il s’agit de Crassula helmsii (T. Kirk) Cockayne, originaire de Nouvelle-Zélande et du sud de l’Australie (Dawson et al., 1987) ; Hydrocotyle ranunculoides L.f., originaire d’Amérique du Nord (Clément et al., 1994); ainsi que Ludwigia grandiflora (Michx.) Greuter & Burdet, Ludwigia peploides (Kunth) P.H. Raven et Myriophyllum aquaticum (Velloso) Verdc., toutes trois originaires d’Amérique du Sud (Raven, 1963 ; Weber, 2003). Ces espèces ont été introduites en Europe par la filière horticole (Vanderhoeven et al., 2011), pour leurs qualités esthétiques et/ou oxygénantes dans les pièces d’eau d’agréments ainsi qu’en aquariophilie. Les premières introductions datent du 19e et du début du 20e siècle pour C. helmsii (Dawson, 1996), M. aquaticum (Thiébaut, 2007), L. grandiflora et L. peploides (Dandelot et al., 2008). Hydrocotyle ranunculoides n’a été observée que plus tardivement, durant les années 1970 (EPPO, 2006). Ces espèces représentent un risque pour la biodiversité et pour le fonctionnement des écosystèmes aquatiques de nos régions. Leurs principaux impacts sont liés à leur capacité à former des tapis denses émergeants (Hussner, 2009 ; Lambert et al., 2010 ; Stiers et al., 2011), modifiant l’apport lumineux au sein du plan d’eau et exerçant une compétition accrue envers les autres végétaux. Certaines de ces espèces jouissent par ailleurs de caractéristiques biologiques qui sont préjudiciables pour les espèces natives, comme la production de substances allélopathiques (Elakovich et al., 1989 ; Dandelot et al., 2008). Les tapis émergents formés par les populations de ces espèces amphibies ont également comme inconvénients de réduire l’usage des plans d’eau pour l’homme (e.g. navigation, pêche, etc.), et d’augmenter les phénomènes de crue par embâcle à l’écoulement (Pot, 2002 ; Hussner et al., 2007).
2Pour ces différentes raisons, les autorités et les gestionnaires de plans d’eaux ont intérêt à limiter l’expansion rapide de ces espèces. Des actions de prévention, combinant règlementations (e.g. Arrêté DEVN0753883A en France, circulaire relative aux espèces exotiques envahissantes en Wallonie), sensibilisation et responsabilisation des acteurs de la filière horticole (e.g. AlterIAS en Belgique, Convenant waterplanten aux Pays-Bas, Horticultural Code of Practice en Grande-Bretagne), sont actuellement mises en place pour limiter les introductions d’espèces amphibies dans de nouveaux plans d’eau. Ces mesures permettent notamment d’empêcher l’essor de nouvelles espèces exotiques et participent à la stratégie de gestion de la problématique à l’échelle régionale et nationale. Néanmoins, la prévention seule ne permet plus d’enrayer les phénomènes d’invasions déjà en cours. Les gestionnaires, pour mettre en place une stratégie de gestion de la problématique complète et cohérente, sont donc également demandeurs de méthodes de lutte efficaces contre les populations existantes. En fonction de l’état d’invasion d’un plan d’eau, ces méthodes doivent leur permettre une éradication totale de l’espèce, ou des mesures d’atténuation ou de confinement permettant d’enrayer la progression des populations (Hulme, 2006).
3Malgré un nombre d’expérimentations grandissant, tant in situ qu’ex situ, il n’est pas évident pour le gestionnaire de savoir vers quel type de lutte s’orienter et d’identifier les bonnes pratiques de lutte permettant de limiter de manière efficace le développement des espèces amphibies invasives. Ceci est notamment dû aux difficultés de comparer les études disponibles du fait de l’utilisation de variables différentes pour juger de l’efficacité des méthodes, ainsi que des différences d’ampleur entre études, tant en termes spatiaux que temporels. S’ajoutent à cela des contraintes légales imposées aux différents gestionnaires, par exemple en termes de doses applicables de biocides. L’objectif de la présente étude est de comparer l’efficacité des différentes méthodes de lutte, en vue d’orienter les gestionnaires vers les méthodes adéquates afin d’éradiquer ou de limiter l’expansion des plantes invasives amphibies.
2. Matériel et méthodes
2.1. Recherche bibliographique
4D’une part, nous avons utilisé les bases de données AGRICOLA, AGRIS, CAB Abstracts, ISI Web of Knowledge, JSTOR, Scirus et SpringerLink pour rechercher les publications se rapportant à des tests de lutte contre les espèces ciblées. D’autre part, les moteurs de recherche Google Scholar, Exalead et Google ont été utilisés pour ajouter aux résultats les manuels et rapports de gestion, présentations de colloques et thèses de doctorat rapportant des tests de méthodes de lutte. Les recherches ont été réalisées pour chaque espèce-cible en associant le nom de l’espèce (noms scientifiques et synonymes, noms vernaculaires français et anglais) aux mots-clés suivants, liés aux méthodes de lutte : active ingredient, barrier, biocontrol, biological, bury*, chaining, chemical, control*, cut*, dredging, dry*, dye, excavat*, eradicat*, flood*, flowdown, froz*, grazing, hand*, harvest*, herbicide, liquid nitrogen, manage*, mechanical, mow, palatability, peroxide hydrogen, physic*, picking, plastic, polythene, pulling, rak*, removal, roto*, shade, tarp*, Truxor. En plus, l’équivalent français de ces mots-clés (le cas échéant), les noms des substances actives connues et les embranchements/classes d’agents de lutte (e.g. insect*) ont également été inclus dans la recherche. Nous avons pris en compte aussi bien les études in situ que les études en mésocosmes, effectuées entre 1969 et 2010. Les études en mésocosmes utilisent communément des réservoirs contenant des quantités d’eau plus ou moins importantes, au sein desquels est disposé un substrat sédimentaire et nutritif. Concernant les études traitant d’agents de lutte biologique, les « mésocosmes » utilisés étaient, le plus souvent, des boites de Petri. Les deux espèces du genre Ludwigia ont été rassemblées car certaines études ne mentionnent que le genre (e.g. UNIMA, 2001).
2.2. Synthèse des informations
5Méthodes de lutte. Selon une typologie classique (Caffrey, 1993), les méthodes de lutte utilisées dans les études retenues ont été classées en plusieurs catégories :
6– « lutte mécanique » : arrachage manuel (Am) et arrachage mécanique (AM) (e.g. moissonneuse aquatique, pelleteuses à godet) ;
7– « lutte chimique » (Ch) ;
8– « lutte biologique » (LB) ;
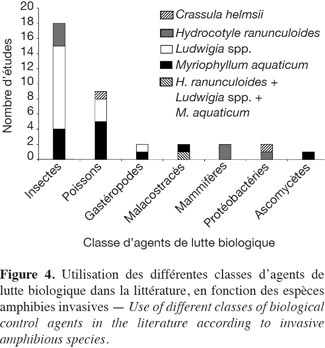
9– « lutte environnementale » (E) telle que l’utilisation de bâches, l’enfouissement ou encore la mise en assec ;
10– les méthodes dites « Autres » (A) reprenant des modalités ne pouvant clairement s’inscrire dans les catégories précitées, telles que l’élévation de la salinité, l’utilisation d’azote liquide/du lance-flamme ou autre traitement thermique (e.g. Waipuna).
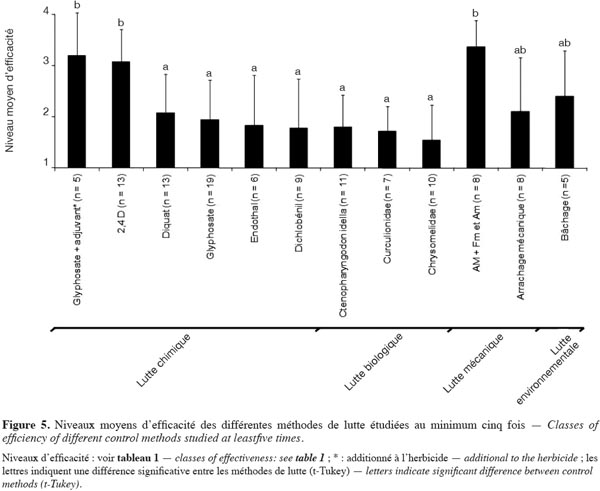
11Pour les luttes mécaniques ou chimiques, nous avons également relevé l’association à des finitions mécaniques (FM) et manuelles (Fm), différentes de la méthode principale. Concernant le traitement chimique, nous nous référons à la substance active et non pas à l’herbicide commercial. L’adjuvant n’est donc pas considéré, sauf si celui-ci est additionné à la formulation de l’herbicide de départ. Les différentes méthodes de lutte sont régulièrement combinées en scénarios :
12– lutte chimique (Ch) combinée à d’autres méthodes de lutte pour la finition (FM, Fm, P) ;
13– lutte mécanique avec arrachages manuel et mécanique utilisés en combinaison ;
14– etc.
15Niveaux d’efficacité. L’efficacité de chaque test de lutte a été évaluée en fonction de la diminution d’abondance des espèces cibles, sur base d’une grille d’évaluation commune (Tableau 1). Selon les études, ces niveaux de diminution sont évalués quantitativement ou qualitativement. Pour les études quantitatives, la variation d’abondance des espèces cibles est chiffrée par la perte de biomasse ou de recouvrement. Pour les études qualitatives, seules des appréciations non chiffrées sont disponibles. Quatre classes de diminution d’abondance ont été prises en compte (Tableau 1). Celles-ci correspondent à la diminution des espèces sur la période considérée dans les études. Par ailleurs, une échelle relative aux études d’appétence en mésocosme, difficilement interprétables en termes d’efficacité in situ, a été développée séparément. Dans celle-ci, seules les correspondances avec des niveaux d’efficacité « faible » ou « modéré » ont été identifiées (Tableau 1).
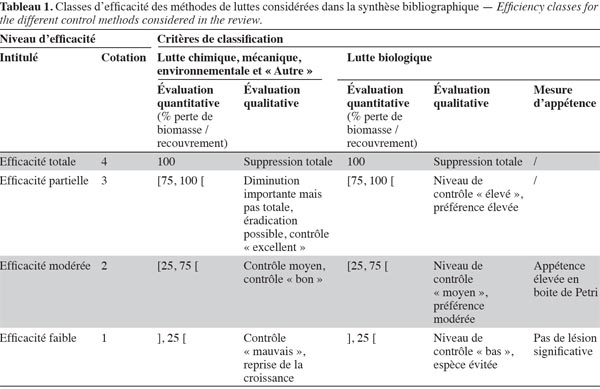
16Modalités d’expérimentation. Pour chaque étude, les informations suivantes ont été notées :
17– tests in situ / en mésocosme ;
18– laps de temps entre l’application de la méthode de lutte et la dernière mesure de son efficacité ;
19– agent de lutte / matière active utilisée (et dosage de la matière active, le cas échéant) ;
20– le nombre de réplications spatiale et/ou temporelle du test.
3. Résultats
21Notre recherche bibliographique a permis de sélectionner 87 sources publiées entre 1969 et 2010, pour l’ensemble des cinq espèces amphibies ciblées. La liste exhaustive de ces références est reprise dans l’Annexe 1.
3.1. Effort de recherche
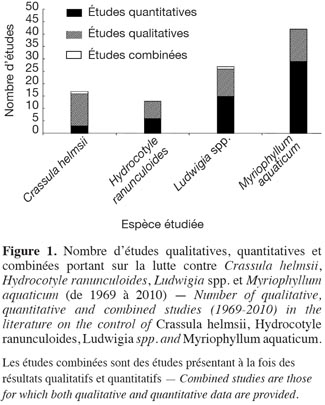
22Comme le montre la figure 1, toutes les espèces n’ont pas fait l’objet du même effort de recherche. Myriophyllum aquaticum est l’espèce qui a fait l’objet du plus grand nombre de tests de lutte (42/87 études, soit 48 %), alors que 28 % des études concernent les deux espèces de Ludwigia, 17 %, C. helmsii et 14 %, H. ranunculoides. Il est à noter qu’une étude peut soit intégrer plusieurs espèces-cibles, soit reprendre des études intégrant à la fois des efficacités qualitatives et quantitatives (« études combinées »). Les études qualitatives sont fortement représentées pour C. helmsii. Les études portant sur M. aquaticum, contrairement, comptent une grande proportion d’études quantitatives.
3.2. Modalités d’expérimentation
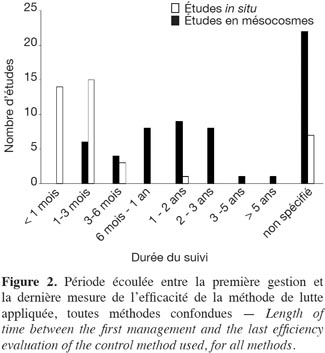
23Pour l’intégralité des études, 56 % des essais ont été effectués in situ contre 44 % en mésocosme. La période entre l’application de la méthode de lutte et la mesure finale de l’efficacité est en général courte : 80 % des études en mésocosme se sont étalées sur une période inférieure à 6 mois (Figure 2). In situ, 2 % des études se sont déroulées sur plus de cinq ans, 36 % sur 1 à 5 ans et 20 % sur moins de 6 mois (Figure 2). Enfin, 18 % des études en mésocosme et 38 % des études in situ ne définissent pas clairement la période entre l’application de la méthode de lutte et la mesure de l’efficacité. Près de la moitié des études ne mentionnent pas si les tests ont été répliqués spatialement. Quarante-et-un pour cent des études mentionnent clairement la considération d’au moins deux réplications spatiales.
3.3. Méthodes de lutte utilisées
24Trente-sept pour cent des études portent sur la lutte chimique, 34 % sur la lutte biologique, 16 % sur la lutte mécanique (AM, AM + Fm, Am) et 13 % sur la lutte environnementale. La combinaison de la lutte chimique avec d’autres méthodes (AM, Fm, mise en assec) est considérée dans 8 % des études. Les méthodes de lutte dites « Autres » sont reprises dans 7 % des études. L’efficacité de 70 % des tests utilisant la lutte chimique a été évaluée quantitativement, de même que 47 % des tests de lutte mécanique, 14 % des tests de lutte biologique et 9 % des tests de lutte environnementale. Quarante pour cent des tests utilisant des agents de lutte biologique intègrent des mesures quantitatives du degré d’appétence en boite de Petri.
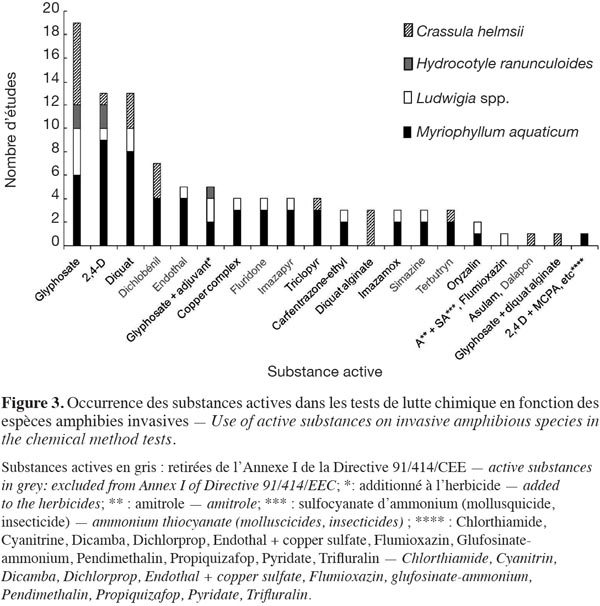
25Concernant la lutte chimique, plusieurs substances actives ont généralement été testées par étude. Les substances actives les plus communément utilisées dans les tests de la lutte chimique (33/87 études) sont le glyphosate (66 % des études chimiques), suivi par le 2,4-D et le diquat se partageant équitablement 60 % des tests de la lutte chimique. Le glyphosate et le 2,4-D ont été testés sur l’ensemble des espèces-cibles (Figure 3). Le dichlobénil a été utilisé dans 21 % des tests de la lutte chimique. D’autres substances actives comme l’endothal, le fluridone, l’imazapyr et le triclopyr ont été considérées. Sur l’ensemble des études, 86 % des substances actives ont été testées quantitativement pour M. aquaticum, 48 % pour Ludwigia spp., 31 % pour C. helmsii et 10 % pour H. ranunculoides. Au niveau européen, plus d’un quart des substances actives ont été retirées de l’annexe I de la Directive 91/414/CEE concernant la mise sur le marché des produits phytopharmaceutiques (Figure 3).
26En lutte biologique, comme le montre la figure 4, les classes des Insectes et des Poissons comprennent les agents de lutte biologique les plus utilisés avec respectivement 58 % et 29 % des études ayant trait à cette méthode de lutte (30 études). La classe des Insectes est représentée par trois familles : les Chrysomelidae (10/18 études), les Curculionidae (7/18 études) et les Libellulidae (1/18 études). Concernant les Poissons, seule la famille des Cyprinidae a été testée, principalement avec deux espèces de carpe : Ctenopharyngodon idella (8/9 études) et Cyprinus carpio (1/9 études). D’autres classes d’agents de lutte biologique sont également représentées (Figure 4).
27Au sein de la lutte mécanique, l’arrachage mécanique a été adopté dans 62 % des études (8/13 études). Suivent l’arrachage mécanique combiné à des finitions manuelles (31 %) et l’arrachage manuel (23 %). Enfin, concernant la lutte environnementale, le bâchage a le plus communément été utilisé (5/11 études).
3.4. Efficacité des méthodes de lutte
28Nous présentons ci-dessous une comparaison globale des méthodes de lutte. Seules les substances actives, agents et méthodes de lutte mécaniques/environnementales ayant été testés par au minimum 5 études sont comparés. Les autres seront abordées dans la discussion. Plusieurs espèces et/ou plusieurs méthodes de lutte ont pu être étudiées dans une même publication scientifique. Dans ce cas, la publication a été considérée comme étant constituée d’autant d’études indépendantes. Un niveau moyen d’efficacité a été calculé sur base d’un système de cotation attribuant des cotes de 1, 2, 3 et 4 pour des niveaux d’efficacité respectivement faible, modéré, partiel et total (Tableau 1). Sur cette base quantitative, une analyse de la variance hiérarchisée à deux facteurs a été réalisée en vue de comparer les méthodes de lutte et les modalités au sein des méthodes de lutte. Celle-ci a montré des différences significatives dans les niveaux moyens d’efficacité, tant entre méthodes de lutte (F3,103 = 7,22 ; P < 0,001) qu’entre les modalités au sein de méthodes de lutte (F8,103 = 5,36 ; P < 0,001). Un test de comparaison des moyennes de Tukey, entre les différentes modalités, a été réalisé. La figure 5 illustre les niveaux moyens d’efficacité des modalités au sein des méthodes de lutte.
29Lutte chimique. Le 2,4-D et le glyphosate combiné avec un adjuvant montrent respectivement un niveau d’efficacité total dans 23 % (15,5 % en mésocosme et 7,5 % in situ) et 40 % des études (20 % en mésocosme et 20 % in situ), alors que le glyphosate utilisé seul ne permet pas d’atteindre ce niveau d’efficacité. Comme le montre la figure 5, au sein de la lutte chimique, le 2,4–D présente une meilleure efficacité que les autres substances actives, hormis le glyphosate combiné avec un adjuvant. Le diquat, le dichlobénil, l’endothal ou le glyphosate (sans adjuvant additionnel) montrent au mieux une efficacité partielle dans 33 % des études leur étant consacrées. Concernant l’utilisation du glyphosate « seul », un seul test appliquant une combinaison mise en assec - glyphosate - finitions manuelles - drainage d’un canal a permis d’atteindre un niveau d’efficacité total sur une population de M. aquaticum (UNIMA, 2001).
30Lutte biologique. Les niveaux d’efficacité des agents de lutte biologique ont majoritairement été étudiés en mésocosme. La figure 5 illustre le niveau faible d’efficacité de la lutte biologique, quel que soit l’agent de lutte utilisé. Seules deux études (in situ) sur 30 utilisant la lutte biologique ont montré une efficacité partielle sur M. aquaticum et sur L. grandiflora, par l’utilisation respectivement de carpes herbivores à forte densité (100 carpes par hectare) et d’une espèce de Chrysomelidae (Lysathia ludoviciana). Dans toutes les autres études effectuées in situ, le niveau d’efficacité atteint avec les autres agents de lutte biologique est modéré à faible.
31Lutte mécanique et environnementale. L’arrachage mécanique combiné à des finitions manuelles (4/87 études) ainsi que l’arrachage manuel seul (2/87 études) montrent des niveaux d’efficacité parmi les meilleurs (Figure 5). Lorsque l’arrachage mécanique est envisagé seul, il ne montre en moyenne pas un tel niveau d’efficacité (Figure 5). Une seule étude qualitative mentionne qu’un curage a permis d’éradiquer C. helmsii. La lutte environnementale (ici utilisation d’une bâche) permet d’atteindre des niveaux d’efficacité partiels sur C. helmsii (Figure 5). Il a été montré, cependant, que cette alternative n’est pas exploitable pour le genre Ludwigia (UNIMA, 2001).
3.5. Cout des méthodes de lutte
32La synthèse bibliographique a montré un manque manifeste d’informations concernant les couts de la lutte sur les différentes espèces. Seules des estimations sont parfois proposées (e.g. £ 10 000·km-1 de linéaire de berge pour l’arrachage mécanique d’H. ranunculoides, Newman et al. [1999] et Pot [2002]). Seules trois études (Dutartre et al., 1993 ; UNIMA, 2001 ; Meisler, 2008) ont donné des estimations de couts sur Ludwigia spp. et M. aquaticum. Ainsi, par ordre croissant de couts de lutte, nous avons : la lutte chimique (21 ± 8,5 € pour 100 m², n = 5), l’arrachage mécanique (92,5 ± 80,5 € pour 100 m², n = 4) et l’arrachage manuel (875 ± 715,5 € pour 100 m², n = 3). Des couts relatifs aux finitions manuelles post-mécanique et post-chimique ont également été considérés. Ils s’élèvent respectivement à 45 et 400 €·m-².
4. Discussion
33Différentes méthodes et scénarios de lutte offrent des niveaux d’efficacité totaux et partiels. Seuls ceux-ci seront examinés plus en détail dans la suite de ce texte, par méthode de lutte. Les tendances générales pour les différentes espèces étant similaires, une approche globale est envisagée en priorité. Néanmoins, des nuances sont apportées entre les espèces, là où des divergences claires existent en termes d’effort de recherche ou de résultats.
4.1. Lutte chimique
34Peu de substances actives ont montré des niveaux d’efficacité totale et ce, malgré un effort de recherche important. Parmi les substances actives ayant donné une efficacité totale, certaines peuvent être écartées par plusieurs restrictions légales (e.g. retrait de l’annexe I de la Directive 91/414/CEE). Les matières actives agréées en Europe qui montrent le plus haut niveau d’efficacité sont le 2,4D et le glyphosate + adjuvant. Pour atteindre un niveau d’efficacité total, plusieurs études soulignent la nécessité de combiner la lutte chimique à d’autres méthodes comme la mise en assec, l’arrachage mécanique, les finitions manuelles ou le drainage (UNIMA, 2001 ; Meisler, 2008 ; Clarke, 2009). Dans ces conditions, il est difficile de quantifier l’efficacité réelle de l’herbicide. Par exemple, une étude sur Ludwigia spp. (Dutartre et al., 1993) a montré qu’après une seule application de 4,32 kgs.a.·ha-1 de glyphosate, la population de Ludwigia sp. revenait quasiment à son stade initial d’invasion après deux années. Pour pallier à ce problème, de nombreuses études ont mis en évidence qu’il était nécessaire de répéter l’application de la substance active plusieurs fois par an et ce, sur plusieurs années (Dawson, 1996 ; Newman et al., 1999 ; Gomes, 2005 ; Hofstra et al., 2006 ; Meisler, 2008 ; Clarke, 2009). In situ, aucun herbicide n’a permis d’atteindre un niveau d’efficacité total sur C. helmsii et H. ranunculoides. En mésocosme, une efficacité totale a été atteinte par application :
35– de triclopyr sur C. helmsii ;
36– de 2,4-D, d’imazapyr ou encore de triclopyr sur M. aquaticum (Negrisoli et al., 2003 ; Hofstra et al., 2006 ; Wersal et al., 2007 ; Champion et al., 2008).
37In situ, seul le 2,4-D a permis d’atteindre ce niveau d’efficacité sur M. aquaticum (Moreira et al., 1999).
38De plus, il n’est plus à démontrer que les herbicides ont un impact non négligeable sur la biodiversité indigène (Tsui et al., 2003 ; Kettenring et al., 2011). L’impact sur la flore native sera plus élevé pour les substances actives ayant un large spectre d’action (e.g. glyphosate, imazapyr), contrairement aux substances actives ayant un spectre d’action limité aux dicotylées (e.g. 2,4-D, triclopyr). Les résultats de la présente synthèse bibliographique soulignent que l’utilisation d’adjuvants (supplémentaire à la formulation de l’herbicide) permet d’augmenter les niveaux d’efficacité (Defra, 2007 ; Newman, 2009). Ces adjuvants ont également une écotoxicité propre (Brausch et al., 2007) et sont soumis aux mêmes obligations en termes d’homologation que les substances actives. Toutefois, il ne faut pas écarter que les autres méthodes de lutte portent, elles aussi, préjudice au milieu géré.
4.2. Lutte biologique
39Comme l’a déjà souligné une étude antérieure sur d’autres espèces invasives (Thomas et al., 2007), l’utilisation d’agents de lutte biologique n’a montré, tout au plus, qu’une efficacité partielle à l’encontre des espèces-cibles. Ce niveau d’efficacité a été obtenu sur M. aquaticum avec Ctenopharyngodon idella, installée dans des plans d’eau à très forte densité (Garner, 2008). Pourtant, d’autres études montrent que l’appétence de l’espèce pour M. aquaticum est moyenne à faible (Pine et al., 1991 ; Catarino et al., 1997 ; Parker et al., 2005). Cet agent de lutte est peu efficace voire inefficace sur les autres espèces-cibles (Dawson et al., 1987 ; Pine et al., 1991 ; Parker et al., 2005). Ctenopharyngodon idella est généraliste et consomme d’autres plantes aquatiques indigènes comme Lemna minor, Ludwigia palustris ou Potamogeton spp. (e.g. Catarino et al., 1997 ; Parker et al., 2005). Concernant L. grandiflora, l’agent de lutte biologique ayant offert un niveau d’efficacité partiel est un Chrysomelidae (Lysathia ludoviciana) (Mc Gregor et al., 1996). Le genre Lysathia offre un niveau d’efficacité modéré sur L. peploides et M. aquaticum (Habeck et al., 1980 ; Cordo et al., 1982 ; Cilliers, 1999).
40Il existe des obstacles non négligeables à l’utilisation d’agents de lutte biologique en Europe. Ainsi, une grande majorité des agents de lutte est originaire des continents américain et asiatique, et représente un risque en termes d’invasion. Certains (e.g. C. idella, Listronotrus sp., Procambarus sp., Ralstonia solanacearum) sont déjà considérés comme espèces invasives dans divers pays européens (EPPO, 2011 ; Verbrugge et al., 2012). Il ne faut néanmoins pas exclure que l’utilisation d’un agent de lutte biologique puisse offrir une solution efficace à l’avenir pour lutter contre certaines espèces.
4.3. Lutte mécanique
41Une éradication de C. helmsii et M. aquaticum a été observée suite à des méthodes simples après une année de recul, respectivement via un seul arrachage mécanique de la couche humique et via un seul arrachage manuel (Leach et al., 2000 ; UNIMA, 2001). Les deux sources mentionnent cependant que ces éradications ne sont vraisemblablement pas généralisables. L’arrachage mécanique, suivi de plusieurs finitions manuelles, a permis d’éradiquer H. ranunculoides et L. grandiflora (Legrand, 2002 ; Newman et al., 2010). Les finitions manuelles ont été effectuées sur deux années pour L. grandiflora. L’application de ce scénario semble relativement simple, mais les réalités de terrain (inaccessibilité des sites pour les engins lourds, difficulté d’observer l’espèce) peuvent fortement en compromettre la faisabilité. Une autre méthode de lutte mécanique a montré un niveau d’efficacité partiel, il s’agit de l’excavation des berges à l’aide d’une déplaqueuse de gazon ou « turf cutter » sur C. helmsii (Clarke, 2009).
4.4. Lutte environnementale
42La mise en assec et l’inondation prolongées (5 à 9 mois) ont montré un niveau d’efficacité total, mais dans des conditions difficilement généralisables (assèchement complet des boues, salinité élevée) (e.g. Legrand, 2002 ; Charlton et al., 2010 ; Wersal, 2010). D’autres méthodes de lutte environnementale ont montré des niveaux d’efficacité partiels. Parmi celles-ci, il y a l’utilisation de barrière benthique (e.g. bâche en polythène) sur C. helmsii (Leach et al., 1999 ; Leach et al., 2000 ; Wilton-Jones, 2005). La barrière benthique est infructueuse sur le genre Ludwigia (Maman et al., 2007).
4.5. Méthodes de lutte dites « Autres »
43La présente synthèse souligne que plusieurs méthodes de lutte alternatives « high-tech » ou « atypique » classées dans la catégorie « Autres » (azote liquide, H2O2, lance-flamme, Waipuna) ont été entreprises, mais n’ont donné que des niveaux d’efficacité modérés sur C. helmsii et M. aquaticum (Bayer et al., 1990 ; Dawson et al., 1991 ; Bridge, 2005). Des recherches plus approfondies sont nécessaires pour connaitre le réel potentiel de ces méthodes.
4.6. Combinaison de méthodes de lutte
44Une étude (UNIMA, 2001) rapporte que la combinaison d’un traitement chimique, d’un arrachage mécanique et d’une finition manuelle est le scénario de gestion donnant un niveau d’efficacité partiel le plus élevé sur Ludwigia spp. Selon la même étude, le même scénario a permis de limiter localement le développement de M. aquaticum.
45Aucune source bibliographique n’intègre la combinaison d’un herbicide et d’un agent biologique pour les espèces-cibles. Pourtant, aux États-Unis, des études en mésocosme effectuées sur d’autres espèces aquatiques ont testé cette combinaison. Elles ont démontré que l’agent de lutte biologique combiné à l’herbicide donnait, à même dose d’herbicide, de meilleurs niveaux d’efficacité que l’herbicide seul (Nelson et al., 2005).
4.7. Discussion générale
46Malgré une forte occurrence dans la littérature, la lutte chimique, potentiellement moins fastidieuse à mettre en place et moins couteuse que les autres méthodes de lutte, n’a montré des niveaux d’efficacité satisfaisants qu’avec certaines matières actives. Dans de nombreux autres cas, les tests de lutte chimique ont montré des résultats relativement peu encourageants. A contrario, malgré leur faible occurrence dans la littérature et leur faible caractère innovant, l’arrachage manuel ou mécanique suivi de finitions manuelles apparait comme une méthode de lutte encourageante. Cette tendance est en accord avec les résultats de Simberloff (2003). Si l’utilisation de certaines substances actives permet d’arriver à des niveaux d’efficacité comparables à ceux obtenus en lutte mécanique, la combinaison de méthodes chimiques et mécaniques de lutte représente certainement une piste de recherche pour l’avenir.
47Au vu des résultats de cette synthèse, plusieurs questions, déjà posées pour les espèces invasives terrestres, peuvent l’être pour les espèces amphibies. Faut-il impérativement « éradiquer » les populations observées ? Faut-il seulement limiter le développement de l’espèce invasive « en apprenant à vivre avec » ? Ou doit-on tout simplement « ne rien faire » ? (Rejmánek et al., 2002 ; Simberloff, 2009). Une éradication pourrait être envisagée pour H. ranunculoides, au contraire de C. helmsii où même l’éradication à un stade initial d’invasion à l’échelle du plan d’eau est laborieuse (Delbart et al., 2011). Pour les autres espèces, la mise en place de mesures d’atténuation semblerait le plus réaliste. Pour empêcher au mieux la propagation de l’espèce à travers les paysages, il est judicieux de cibler les plans d’eau/populations à risque (Wadsworth et al., 2000), c’est-à-dire les plans d’eau envahis étant à la source de nouvelles populations-satellites (e.g. plan d’eau en amont de bassin versant ; plan d’eau abritant des espèces animales pouvant disperser des fragments de plantes ; plan d’eau faisant l’objet de curage ; etc.). Des priorités de gestion doivent donc être dégagées dans ce sens. À côté de ces difficultés de lutte, il ne faut pas exclure l’existence de lacunes sur le degré réel d’occurrence de ces espèces à l’échelle régionale et sur leurs capacités de dispersion (Hussner et al., 2010 ; Delbart et al., 2011). Ces connaissances sont nécessaires pour garantir le succès d’une potentielle éradication (Myers et al., 2000). Enfin, plusieurs contraintes peuvent par ailleurs compliquer la gestion. Parmi celles-ci, citons : la cohabitation de plusieurs espèces invasives aquatiques au sein d’un même plan d’eau ; la nécessité d’adapter la méthode de lutte (e.g. arrachage manuel vs mécanique) en fonction du substrat (Lambert et al., 2010) ; la présence d’espèces d’intérêt patrimonial ; la présence d’obstacles (e.g. arbres morts, pollution des vases) ou encore la ré-invasion potentielle par une autre plante aquatique invasive (e.g. recolonisation par Elodea nuttallii en post-gestion de C. helmsii) (Reid et al., 2009 ; Delbart, observation personnelle).
48L’analyse des études sur les méthodes de lutte, réalisée dans le présent travail, s’est parfois heurtée à un manque d’informations dans les publications prises en compte. Ainsi, par exemple, il n’était pas toujours possible de connaitre le niveau initial d’invasion ou la taille des populations avant application des méthodes de lutte, ni de savoir si des actions de lutte avaient été entreprises antérieurement. Ces données peuvent cependant se révéler essentielles pour comparer et mesurer l’efficacité des actions de lutte engagées.
49À l’issue de la présente synthèse bibliographique, plusieurs recommandations peuvent être formulées en vue d’améliorer les connaissances sur les moyens de lutte contre les plantes amphibies invasives :
50– des recherches quantitatives et répliquées spatialement sont encore nécessaires pour mieux évaluer le succès et les couts des méthodes de lutte mécanique ;
51– les études évaluant le succès de méthodes de lutte devraient systématiquement inclure des populations témoins, vu la variabilité interannuelle des taux d’invasion parfois observée ;
52– la combinaison de méthodes (e.g. traitement chimique avec arrachage manuel en finition ; arrachage mécanique combiné à la lutte biologique, etc.) représente une piste de recherche pour l’avenir.
53Remerciements
54Nous tenons à remercier Gwenn Frisson pour sa relecture avisée ainsi qu’Alain Dutartre pour sa collaboration concernant l’obtention de certains ouvrages. Nos remerciements vont également au Service public de Wallonie (DCENN, MM. Stéphan Adant et Francis Lambot) pour la confiance qu’ils nous ont témoignée durant nos recherches.
Bibliographie
Bayer D.E. & Rejmánková E., 1990. Removal of herbicide residua and nitrates from agricultural waters by aquatic plants. WRC Project W-727. Berkeley, CA, USA: University of California, Water Resources Center.
Branquart E., 2009. Guidelines (version 2.6) for environmental impact assessment and list classification of non-native organisms in Belgium. Brussels: Belgian Forum on Invasive Species.
Brausch J., Beall B. & Smith P., 2007. Acute and sub-lethal toxicity of three POEA surfactant formulations to Daphnia magna. Bull. Environ. Contam. Toxicol., 78, 510-514.
Bridge T., 2005. Controlling New Zealand pygmyweed Crassula helmsii using hot foam, herbicide and by burying at Old Moor RSPB Reserve, South Yorkshire, England. Conserv. Evidence, 2, 33-34.
Caffrey J., 1993. Plant management as an integrated part of Ireland’s aquatic resources. Hydroécologie Appliquée, 5, 77-96.
Catarino L.F., Ferreira M.T. & Moreira I.S., 1997. Preferences of grass carp for macrophytes in Iberian drainage channels. J. Aquat. Plant Manage., 36, 79-83.
Champion P.D., James T.K. & Carney E.C., 2008. Safety of Triclopyr triethylamine to native wetland species. New Zealand Plant Prot., 61, 378-383.
Charlton P.E., Gurney M. & Graeme Lyons G., 2010. Large-scale eradication of New Zealand pygmyweed Crassula helmsii from grazing marsh by inundation with seawater, Old Hall Marshes RSPB reserve, Essex, England. Conserv. Evidence, 7, 130-133.
Cilliers C.J., 1999. Lysathia n.sp. (Coleoptera: Chrysomelidae), a host-specific beetle for the control of the aquatic weed Myriophyllum aquaticum (Haloragaceae) in South Africa. Hydrobiologia, 415, 271-276.
Clarke S., 2009. A summary of three different approaches to the treatment of non-native invasive species Crassula helmsii at protected sites. In: Newman J., ed. Abstracts and proceedings of the 41st Robson meeting, 17-18 February 2009, Centre for Ecoloy and Hydrology, Wallingford, UK. Wallingford, UK: Centre for Ecoloy and Hydrology, 14-17.
Clément E.J. & Foster M.C., 1994. Alien plants of the British Isles. London: Botanical Society of the British Isles.
Cordo H.A. & DeLoach C.J., 1982. The flea beetle, Lysathia flavipes, that attacks Ludwigia (water primrose) and Myriophyllum (parrot feather) in Argentina. Coleopterists Bull., 36, 298-301.
Dandelot S. et al., 2008. Allelopathic potential of two invasive alien Ludwigia spp. Aquat. Bot., 88, 311-316.
Dawson F.H., 1996. Crassula helmsii: attempts at elimination using herbicides. Hydrobiologia, 340, 241-245.
Dawson F.H. & Warman E.A., 1987. Crassula helmsii (T.Kirk) Cockayne: is it an aggressive alien aquatic in Britain? Environ. Conserv., 42, 247-272.
Dawson F.H. & Henville P., 1991. An investigation of the control of Crassula helmsii by herbicidal chemicals (with interim guidelines on control). Final Report. Peterborough, UK: Nature Conservancy Council.
DEFRA (Department for Environment, Food & Rural Affairs), 2007. Research project final report: development of eradication strategies for Ludwigia species. London: DEFRA, http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More&Location=None&Completed=1&ProjectID=14763, (21.01.2011).
Delbart E., Monty A. & Mahy G., 2011. Gestion de Crassula helmsii en Belgique plus difficile qu’il n’y parait ? EPPO/OEPP Bull., 41, 226-231.
Dutartre A. & Oyarzabal J., 1993. Gestion des plantes aquatiques dans les lacs et les étangs landais. Hydroécologie appliquée, 5(2), 43-60.
Elakovich S.D. & Wooten J.W., 1989. Allelopathic potential of sixteen aquatic and wetland plants. J. Aquat. Plant Manage., 27, 78-84.
EPPO (European and Mediterranean Plant Protection Organization), 2006. Data sheets on quarantine pests: Hydrocotyle ranunculoides. EPPO/OEPP Bull., 36, 3-6.
EPPO (European and Mediterranean Plant Protection Organization), 2011. EPPO A1 and EPPO A2 lists of pests recommend for regulation as quarantaine pests. Paris : EPPO, http://archives.eppo.org/EPPOStandards/PM1_GENERAL/pm1-02%2820%29_A1A2_2011.pdf, (12.01.2012).
Garner A.B., 2008. High-density grass carp stocking effects on a reservoir invasive plant, water quality, and native fishes. Raleigh, NC, USA: North Carolina State University.
Gomes B., 2005. Controlling New Zealand pygmyweed Crassula helmsii in field ditches and a gravel pit by herbicide spraying at Dungeness RSPB Reserve, Kent, England. Conserv. Evidence, 2, 62.
Habeck D.H. & Wilkerson R., 1980. The life cycle of Lysathia ludoviciana (Fall) (Coleoptera: Chrysomelidae) on parrotfeather, Myriophyllum aquaticum (Velloso) Verde. Coleopterists Bull., 34, 167-170.
Hofstra D.E., Champion P.D. & Dugdale T.M., 2006. Herbicide trials for the control of parrotsfeather. J. Aquat. Plant Manage., 44, 13-18.
Hulme P.E., 2006. Beyond control: wider implications for the management of biological invasions. J. Appl. Ecol., 43, 835-847.
Hussner A., 2009. Growth and photosynthesis of four invasive aquatic plant species in Europe. Weed Res., 49, 506-515.
Hussner A. & Lösch R., 2007. Growth and photosynthesis of Hydrocotyle ranunculoides L. fil. in Central Europe. Flora, 202, 653-660.
Hussner A., Van de Weyer K., Gross E.M. & Hilt S., 2010. Comments on increasing number and abundance of non-indigenous aquatic macrophyte species in Germany. Weed Res., 50, 519-526.
Kettenring K.M. & Reinhardt Adams C., 2011. Lessons learned from invasive plant control experiments: a systematic review and meta-analysis. J. Appl. Ecol., 48, 970-979.
Lambert E., Dutartre A., Coudreuse J. & Haury J., 2010. Relationships between the biomass production of invasive Ludwigia species and physical properties of habitats in France. Hydrobiologia, 656, 173-186.
Leach J. & Dawson H., 1999. Crassula helmsii in British Isles – an unwelcome invader. Br. Wildl., 10, 234-239.
Leach J. & Dawson H., 2000. Is resistance futile? The battle against Crassula helmsii. J. Pract. Ecol. Conserv., 4, 7-17.
Legrand C., 2002. Pour contrôler la prolifération des jussies (Ludwigia spp.) dans les zones humides méditerranéennes. Guide technique. Montpellier, France : Agence Méditerranéenne de l’Environnement.
Maman L. & Hurtrez L., 2007. Bilan des retours d’expériences sur les espèces envahissantes du Bassin Loire Bretagne et recommandations de gestion. Agence de l'eau Loire-Bretagne, http://centrederessources-loirenature.com/mediatheque/especes_inva/telechargements/evenementiel/expose_maman_09_03_07.pdf, (25.02.2010).
Mc Gregor S.A. et al., 1996. The potential for biological control of water primrose (Ludwigia grandiflora) by the water primrose FleaBeetle (Lysathia ludoviciana) in the Southeastern United States. J. Aquat. Plant Manage., 34, 74-76.
Meisler J., 2008. Ludwigia control project: final report. Laguna de Santa Rosa, Sonoma County, California. Santa Rosa, CA, USA: Laguna de Santa Rosa Foundation, http://www.lagunafoundation.org/pdfs/Ludwigia%20Control%20Project%20Final%20Report.pdf, (25.06.2011).
Moreira I., Monteira A. & Ferreira T., 1999. Biology and control of parrotfeather (Myriophyllum aquaticum) in Portugal. Ecol. Environ. Conserv., 5, 171-179.
Myers J.H., Simberloff D., Kuris A.M. & Carey J.R., 2000. Eradication revisited: dealing with exotic species. Trends Ecol. Evol., 15, 316-320.
Negrisoli E. et al., 2003. Uso de diferentes herbicidas no controle de Myriophyllum aquaticum. Planta Daninha, 21, 89-92.
Nelson L.S. & Shearer J.F., 2005. 2,4-D and Mycoleptodiscus terrestris for control of Eurasian watermilfoil. J. Aquat. Plant Manage., 43, 29-34.
Newman J.R., 2009. Topfilm the only approved adjuvant for aquatic use and its value. In: Newman J., ed. Abstracts and proceedings of the 41st Robson meeting, 17-18 February 2009, Centre for Ecoloy and Hydrology, Wallingford, UK. Wallingford, UK: Centre for Ecoloy and Hydrology, 20-21.
Newman J.R. & Dawson F.H., 1999. Ecology, distribution and chemical control of Hydrocotyle ranunculoides in the UK. Hydrobiologia, 415, 295-298.
Newman J.R. & Duenas S.A., 2010. Aquatic plant management group, information sheet: control of floating pennywort (Hydrocotyle ranunculoides), http://www.ceh.ac.uk/sci_programmes/documents/Hydrocotyleranunculoides.pdf, (18.01.2011).
Parker J.D. & Hay M.E., 2005. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol. Lett., 8, 959-967.
Pine R.T. & Anderson L.W.J., 1991. Plant preference of triploid grass carp. J. Aquat. Plant Manage., 29, 80-82.
Pot R., 2002. Invasion and management of Floating Pennywort (Hydrocotyle ranunculoides L.f.) and some other alien species in The Netherlands. In: Proceedings of the 11th EWRS International symposium on aquatic weeds, September 2-6 2002, Moliets et Maâ, France. Doorwerth, The Netherlands: European Weed Research Society (EWRS), 435-438.
Raven P.H., 1963. The old world species of Ludwigia (including Jussiae), with a synopsis of the genus (Onagraceae). Reinwardtia, 6, 327-427.
Reid A.M. et al., 2009. Does invasive plant management aid the restoration of natural ecosystems? Biol. Conserv., 142, 2342-2349.
Rejmánek M. & Pitcairn M.J., 2002. When is eradication of exotic pest plants a realistic goal? In: Veitch C.R. & Clout M.N., eds. Turning the tide: the eradication of invasive species. Gland, Switzerland: IUCN Species Survival Commission, 249-253.
Simberloff D., 2003. How much information on population biology is needed to manage introduced species? Conserv. Biol., 17, 83-92.
Simberloff D., 2009. We can eliminate invasions or live with them: successful management projects. Biol. Invasions, 11, 149-157.
Stiers I., Crohain N., Josens G. & Triest L., 2011. Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biol. Invasions, 13, 2715-2726.
Thiébaut G., 2007. Invasion success of non-indigenous aquatic and semi-aquatic plants in their native and introduced ranges. A comparison between their invasiveness in North America and France. Biol. Invasions, 9, 1-12.
Thomas M.B. & Reid A.M., 2007. Are exotic natural enemies an effective way of controlling invasive plants? Trends Ecol. Evol., 22, 447-453.
Tsui M.T.K. & Chu L.M., 2003. Aquatic toxicity of glyphosate-based formulations: comparison between different organisms and the effects of environmental factors. Chemosphere, 12, 1189-1197.
UNIMA, 2001. Expérimentations de techniques de contrôle de Ludwigia sp. et de Myriophyllum brasiliense sur les marais littoraux de Charente-Maritime. La Rochelle, France : Conseil Général de la Charente-Maritime.
Vanderhoeven S. et al., 2011. Perception and understanding of invasive alien species issues by nature conservation and horticulture professionals in Belgium. Environ. Manage., 47, 425-442.
Verbrugge L.N.H. et al., 2012. Risk classifications of aquatic non-native species: application of contemporary European assessment protocols in different biogeographical settings. Aquat. Invasions, 7(1), 49-58.
Wadsworth R.A. et al., 2000. Simulating the spread and management of alien riparian weeds: are they out of control? J. Appl. Ecol., 37, 28-38.
Weber E., 2003. Invasive plant species of the world. A reference guide to environmental weeds. London: CABI Publishing.
Wersal R.M., 2010. The conceptual ecology and management of parrotfeather [Myriophyllum aquaticum (Vell.) Verdc.]. Starkville, MS, USA: Mississippi State University.
Wersal R.M. & Masden J.D., 2007. Comparison of imazapyr and imazamox for control of parrotfeather (Myriophyllum aquaticum (Vell.) Verdc.). J. Aquat. Plant Manage., 45, 132-136.
Wilton-Jones G., 2005. Control of New Zealand pygmyweed Crassula helmsii by covering with black polythene at The Lodge RSPB Reserve, Bedforshire, England. Conserv. Evidence, 2, 63.
Annexe I. Liste exhaustive des études considérées dans la synthèse bibliographique.
Barreto R., Charudattan R., Pomella A. & Hanada R., 2000. Biological control of neotropical aquatic weeds with fungi. Crop Prot., 19, 697-703.
Bayer D.E. & Rejmánková E., 1990. Removal of herbicide residua and nitrates from agricultural waters by aquatic plants. WRC Project W-727. Berkeley, CA, USA: University of California, Water Resources Center.
Bernhardt E.A. & Duniway J.M., 1984. Root and stem rot of parrotfeather (Myriophyllum brasiliense) caused by Pythium carolinianum. Plant Dis., 68, 999-1003.
Bridge T., 2005. Controlling New Zealand pygmyweed Crassula helmsii using hot foam, herbicide and by burying at Old Moor RSPB Reserve, South Yorkshire, England. Conserv. Evidence, 2, 33-34.
Brunson M.W., 1998. Aquatic weed control: using grass carp. Starkville, MS, USA: Mississippi State University Extension Service, southernsportsmanaquaticsandland.com/yahoo_site_admin/assets/docs/Weeds_Controled_by_Grass_Carp.13761908.pdf, (07.02.2010).
Catarino L.F., Ferreira M.T. & Moreira I.S., 1997. Preferences of grass carp for macrophytes in Iberian drainage channels. J. Aquat. Plant Manage., 36, 79-83.
Champion P.D., James T.K. & Carney E.C., 2008. Safety of triclopyr triethylamine to native. New Zealand Plant Prot., 61, 378-383.
Charlton P.E., Gurney M. & Graeme Lyons G., 2010. Large-scale eradication of New Zealand pygmyweed Crassula helmsii from grazing marsh by inundation with seawater, Old Hall Marshes RSPB reserve, Essex, England. Conserv. Evidence, 7, 130-133.
Child L. & Spencer-Jones D., 1995. Treatment of Crassula helmsii - a case study. In: Pyšek P., Prach K., Rejmánek M. & Wade M., eds. Plant invasions - general aspects and special problems. Amsterdam, The Netherlands: SBP Academic Publ., 195-202.
Cilliers C.J., 1999. Lysathia n.sp. (Coleoptera: Chrysomelidae), a host-specific beetle for the control of the aquatic weed Myriophyllum aquaticum (Haloragaceae) in South Africa. Hydrobiologia, 415, 271-276.
Clark W.E., 1976. Notes on the life history and habits of Perigaster cretura (Herbst) (Coleoptera: Curculionidae) with descriptions of the larva and pupa. Coleopterists Bull., 30, 159-165.
Clarke S., 2009. A summary of three different approaches to the treatment of non-native invasive species Crassula helmsii at protected sites. In: Newman J., ed. Abstracts and proceedings of the 41st Robson meeting, 17-18 February 2009, Centre for Ecoloy and Hydrology, Wallingford, UK. Wallingford, UK: Centre for Ecoloy and Hydrology, 14-17.
Cordo H.A., DeLoach C.J. & Ferrer R., 1981. Biological studies on two weevils, Ochetina bruchi and Onychylis cretatus, collected from Pistia and other aquatic plants in Argentina. Ann. Entomol. Soc. Am., 74, 363-368.
Cordo H.A. & Deloach C.J., 1982a. Notes on the weevils Tyloderma, Auleutes, and Onychylis that feed on Ludwigia and other aquatic plants in Southern South America. Coleopterists Bull., 36, 291-297.
Cordo H.A. & DeLoach C.J., 1982b. The flea beetle, Lysathia flavipes, that attacks Ludwigia (water primrose) and Myriophyllum (parrot feather) in Argentina. Coleopterists Bull., 36, 298-301.
Cordo H.A. & Deloach C.J., 1982c. Weevils Listronotus marginicollis and L. cinnamomeus that feed on Limnobium and Myriophyllum in Argentina. Coleopterists Bull., 36, 302-308.
Cordo H.A., DeLoach C.J. & Ferrer R., 1982. The weevils Lixellus, Tanysphiroideus and Cyrtobagous that feed on Hydrocotyle and Salvinia in Argentina. Coleopterists Bull., 36, 279-286.
Dandelot S., Ferretti S. & Abou-Hamdan H., 2004. Analyse de l’efficacité d’un traitement par le sel pour lutter contre la prolifération de la Jussie Ludwigia peploides Kunth (Raven) en Camargue. Aix-en-Provence, France : Parc naturel régional de Camargue, Université Aix-Marseille III.
Dauphin P., 1996. Les Ludwigia (Oenotheracées), plante-hôtes des Galerucella du groupe Nympheae (Col. Chrysomelidae). Bull. Soc. Linn. Bordeaux, 24(1), 49-50.
Dawson F.H., 1994. Spread of Crassula helmsii in Britain. In: de Waal et al., eds. Ecology and management of invasive riverside plants. Chichester, UK: John Wiley and Sons, 1-14.
Dawson F.H., 1996. Crassula helmsii: attempts at elimination using herbicides. Hydrobiologia, 340, 241-245.
Dawson F.H., 1999. Crassula helmsii - Focus on control. English Nature, Institute of Freshwater Ecology and Institute of Terrestrial Ecology.
Dawson F.H. & Warman E.A., 1987. Crassula helmsii (T.Kirk) Cockayne: is it an aggressive alien aquatic in Britain? Environ. Conserv., 42, 247-272.
Dawson F.H. & Henville P., 1991. An investigation of the control of Crassula helmsii by herbicidal chemicals (with interim guidelines on control). Final Report. Peterborough, UK: Nature Conservancy Council.
DEFRA (Department for Environment, Food & Rural Affairs), 2007. Research project final report: development of eradication strategies for Ludwigia species. London: DEFRA, http://randd.defra.gov.uk/Default.aspx?Menu=Menu&Module=More&Location=None&Completed=1&ProjectID=14763, (21.01.2011).
Dutartre A. & Oyarzabal J., 1993. Gestion des plantes aquatiques dans les lacs et les étangs landais. Hydroécologie appliquée, 5(2), 43-60.
Emerine S.E. et al., 2010. Greenhouse response of six aquatic invasive weeds to imazamox. J. Aquat. Plant Manage., 48, 105-111.
Garner A.B., 2008. High-density grass carp stocking effects on a reservoir invasive plant, water quality, and native fishes. Raleigh, NC, USA: North Carolina State University.
Genovesi P., 2005. Eradications of invasive alien species in Europe: a review. Biol. Invasions, 7, 127-133.
Glomski L.A.M., Poovey A.G. & Getsinger K.D., 2006. Effect of carfentrazone-ethyl on three submersed macrophytes. J. Aquat. Plant Manage., 44, 67-69.
Gomes B., 2005. Controlling New Zealand pygmyweed Crassula helmsii in field ditches and a gravel pit by herbicide spraying at Dungeness RSPB Reserve, Kent, England. Conserv. Evidence, 2, 62.
Gray C.J., Madsen J.D., Wersal R.M. & Getsinger K.D., 2007. Eurasian watermilfoil and parrotfeather control using carfentrazone-ethyl. J. Aquat. Plant Manage., 45, 43-46.
Habeck D.H. & Wilkerson R., 1980. The life cycle of (Fall) Lysathia ludoviciana (Coleoptera: Chrysomelidae) on parrotfeather, Myriophyllum aquaticum (Velloso) Verde. Coleopterists Bull., 34, 167-170.
Haller W.T., Sutton D.L. & Barlowe W.C., 1974. Effects of salinity on growth of several aquatic macrophytes. Ecology, 55, 891-894.
Hofstra D.E., Champion P.D. & Dugdale T.M., 2006. Herbicide trials for the control of parrotsfeather. J. Aquat. Plant Manage., 44, 13-18.
Hong J.C. et al., 2008. Detection of Ralstonia solanacearum in irrigation ponds and aquatic weeds associated with the ponds in North Florida. Plant Dis., 92, 1674-1682.
Hussner A. & Lösch R., 2007. Growth and photosynthesis of Hydrocotyle ranunculoides L. fil. in Central Europe. Flora, 202, 653-660.
Jacot-Guillarmod A., 1977. Myriophyllum, an increasing water weed menace for South Africa. South Afr. J. Sci., 73, 89-90.
Kelly A., 2006. Removal of invasive floating pennywort Hydrocotyle ranunculoides from Gillingham Marshes, Suffolk, England. Conserv. Evidence, 3, 52-53.
Ki Wong P.K., Liang Y., Liu N.Y. & Qiu JW., 2010. Palatability of macrophytes to the invasive freshwater snail Pomacea canaliculata: differential effects of multiple plant traits. Freshwater Biol., 55, 2023-2031.
Leach J. & Dawson H., 1999. Crassula helmsii in British Isles – an unwelcome invader. Br. Wildl., 10, 234-239.
Leach J. & Dawson H., 2000. Is resistance futile? The battle against Crassula helmsii. J. Pract. Ecol. Conserv., 4, 7-17.
Legrand C., 2002. Guide technique : pour contrôler la prolifération des jussies (Ludwigia spp.) des zones humides méditerranéennes. Montpellier, France : Agence Méditerranéenne de l’Environnement.
Machado C. & Rocha F., 1998. Control of Myriophyllum aquaticum in drainage and irrigated channels of the Mondego river valley, Portugal. In: Monteiro A., Vasconcelos T. & Catarino L., eds. Management and ecology of aquatic weeds. Proceedings of the 10th EWRS International Symposium on Aquatic Weeds, 21-25 September 1998, Lisbon, Portugal. Lisboa: Associação Portuguesa de Recursos Hídricos, 373-375.
Maltchik L., Rolon A.S. & Schott P., 2007. Effects of hydrological variation on the aquatic plant community in a floodplain palustrine wetland of southern Brazil. Limnology, 8, 23-28.
Maman L. & Hurtrez L., 2007. Bilan des retours d’expériences sur les espèces envahissantes du Bassin Loire Bretagne et recommandations de gestion. Agence de l'eau Loire-Bretagne, http://centrederessources-loirenature.com/mediatheque/especes_inva/telechargements/evenementiel/expose_maman_09_03_07.pdf, (25.02.2010).
Masser M.P., Murphy T.R. & Shelton J.L., 2001. Aquatic weed management: herbicides. SRAC Publ., 361.
Mc Gregor S.A. et al., 1996. The potential for biological control of water primrose (Ludwigia grandiflora) by the water primrose FleaBeetle (Lysathia ludoviciana) in the Southeastern United States. J. Aquat. Plant Manage., 34, 74-76.
Meisler J., 2008. Ludwigia control project: final report. Laguna de Santa Rosa, Sonoma County, California. Santa Rosa, CA, USA: Laguna de Santa Rosa Foundation, http://www.lagunafoundation.org/pdfs/Ludwigia%20Control%20Project%20Final%20Report.pdf, (25.06.2011).
Monteiro A. & Moreira I., 1990. Chemical control of parrotfeather (Myriophyllum aquaticum). In: Proceedings of the 8th EWRS International Symposium on Aquatic Weeds. Lisbon, Portugal: Instituto Superior de Agronomia, 163-164.
Moreira I., Ferreira T. & Monteiro A., 1989. Aquatic weed bioecology and control in Portugal: a review. In: Bianchi A., ed. Portuguese-German cooperation in applied agricultural results. Vila Reale, Portugal: Universidade de Vila Real, 71-106.
Moreira I., Monteira A. & Ferreira T., 1999. Biology and control of parrotfeather (Myriophyllum aquaticum) in Portugal. Ecol. Environ. Conserv., 5, 171-179.
Morris M.J., Wood A.R. & den Breeÿen A., 1999. Plant pathogens and biological control of weeds in South Africa: a review of projects and progress during the last decade. Afr. Entomol. Mem., 1, 129-137.
Murphy J.E. et al., 2002. Toxic and feeding deterrent effects of native aquatic macrophytes on exotic grass carp (Ctenopharyngodon idella). Ecotoxicology, 9, 243-254.
Napompeth B., 1994. Biological control of paddy and aquatic weeds in Thailand. Bangkok: National Biological Control Research Center (NBCRC), Kasetsart University, www.agnet.org/library/bc/45011/bc45011.pdf, (24.01.2011).
Nayek T.K. & Banerjee T.C., 1987. Life history and host specificity of Altica cyanea (Coleoptera: Chrysomelidae), a potential biological control agent for water primrose, Ludwigia adscendens. Entomophaga, 32, 407-414.
Negrisoli E. et al., 2003. Uso de diferentes herbicidas no controle de Myriophyllum aquaticum. Planta Daninha, 21, 89-92.
Newman J.R., 2009. Topfilm the only approved adjuvant for aquatic use and its value. In: Newman J., ed. Abstracts and proceedings of the 41st Robson meeting, 17-18 February 2009, Centre for Ecoloy and Hydrology, Wallingford, UK. Wallingford, UK: Centre for Ecoloy and Hydrology, 20-21.
Newman J.R. & Dawson F.H., 1999. Ecology, distribution and chemical control of Hydrocotyle ranunculoides in the UK. Hydrobiologia, 415, 295-298.
Newman J.R. & Duenas S.A., 2010. Aquatic plant management group, information sheet: control of floating pennywort (Hydrocotyle ranunculoides), http://www.ceh.ac.uk/sci_programmes/documents/Hydrocotyleranunculoides.pdf, (18.01.2011).
Oberholzer I.G., Mafokoane D.L. & Hill M.P., 2007. The biology and laboratory host range of the weevil, Listronotus marginicollis (Hustache) (Coleoptera: Curculionidae), a natural enemy of the invasive aquatic weed Myriophyllum aquaticum (Velloso) Verde (Haloragaceae) (Parrot's Feather). Afr. Entomol., 15, 385-390.
Oreska M.P.J. & Aldridge D.C., 2011. Estimating the financial costs of freshwater invasive species in Great Britain: a standardized approach to invasive species costing. Biol. Invasions, 13, 305-319.
Parker J.D. & Hay M.E. 2005. Biotic resistance to plant invasions? Native herbivores prefer non-native plants. Ecol. Lett., 8, 959-967.
Petelczyc M., Dutartre A. & Dauphin P., 2006. La jussie (Ludwigia grandiflora) plante hôte d'Altica lythri Aubé (Coleoptera Chrysomelidae), observation in situ dans la Réserve Naturelle du Marais d'Orx (Landes) et en laboratoire. Bull. Soc. Linn. Bordeaux, 34(3), 221-228.
Pine R.T. & Anderson L.W.J., 1991. Plant preference of triploid grass carp. J. Aquat. Plant Manage., 29, 80-82.
Pipet N., 2007. Maîtrise de la colonisation et de la prolifération des jussies dans le marais Poitevin. Travail de synthèse.
Pot R., 2002. Invasion and management of Floating Pennywort (Hydrocotyle ranunculoides L.f.) and some other alien species in The Netherlands. In: Proceedings of the 11th EWRS International symposium on aquatic weeds, September 2-6 2002, Moliets et Maâ, France. Doorwerth, The Netherlands: European Weed Research Society (EWRS), 435-438.
Rejmánková E., 1992. Ecology of creeping macrophytes with special reference to Ludwigia peploides Raven. Aquat. Bot., 43, 283-299.
Richardson R.J. et al., 2008. Response of selected aquatic invasive weeds to flumioxazin and carfentrazone-ethyl. J. Aquat. Plant Manage., 46, 154-158.
Ruiz-Avila R.J. & Klemm V.V., 1996. Management of Hydrocotyle ranunculoides L.f., an aquatic invasive weed of urban waterways in Western Australia. Hydrobiologia, 340, 187-190.
Sabbatini M.R., Murphy K.J. & Irigoyen J.H., 1998. Vegetation-environment relationships in irrigation channel systems of southern Argentina. Aquat. Bot., 60, 119-133.
Sikka H.C., Lynch R.S. & Lindenberger M., 1974. Uptake and metabolism of dichlobenil by emersed aquatic plants. J. Agric. Food Chem., 22, 230-234.
Sisitka, 1998. comm pers. in Leach J. & Dawson H., 1999. Crassula helmsii in British Isles – an unwelcome invader. Br. Wildl., 10, 234-239.
Spencer-Jones D., 1994. Some observations on the use of herbicides for control of Crassula helmsii. In: de Waal L.C., Child L.E., Wade P.M. & Brock J.H., eds. Ecology and management of invasive riverside plants. Chichester, UK: John Wiley and Sons, 15-18.
Sutton D.L. & Bingham S.W., 1969. Absorption and translocation of simazine in parrotfeather. Weed Sci., 17, 431-435.
Sutton D.L. & Bingham S.W., 1970. Uptake of diquat in parrotfeather. Hyacinth Control J., 8, 2-4.
Sutton D.L., Bingham S.W. & Barlowe W.C., 1969. Responses of aquatic plants to combinations of endothall and copper. Weed Sci., 19, 643-646.
Sutton D.L. & Blackburn R.D., 1971. Uptake of copper by parrotfeather. Weed Sci., 19, 282-285.
Thiébaut G. & Gierlinski P., 2007. Gammarid (Crustacea: Amphipoda) herbivory on native and alien freshwater macrophytes. In: Tokarska-Guzik B. et al., eds. Plant invasions: human perception, ecological impacts and management. Leiden, The Netherlands: Backhuys Publishers, 333-340.
Turgut C., 2007. The impact of pesticides toward parrotfeather when applied at the predicted environmental concentration. Chemosphere, 66, 469-473.
UNIMA, 2001. Expérimentations de techniques de contrôle de Ludwigia sp. et de Myriophyllum brasiliense sur les marais littoraux de Charente-Maritime. La Rochelle, France : Conseil Général de la Charente-Maritime.
Washington State Noxious Weed Control Board, 2011. Parrotfeather. Myriophyllum aquaticum, http://www.nwcb.wa.gov/detail.asp?weed=94, (18.12.2012).
Wersal R.M., 2010. The conceptual ecology and management of parrotfeather [Myriophyllum aquaticum (Vell.) Verdc.]. Starkville, MS, USA: Mississippi State University.
Wersal R.M. & Masden J.D., 2007. Comparison of imazapyr and imazamox for control of parrotfeather (Myriophyllum aquaticum (Vell.) Verdc.). J. Aquat. Plant Manage., 45, 132-136.
Wersal R.M. & Masden J.D., 2010. Comparison of subsurface and foliar herbicide applications for control of parrotfeather (Myriophyllum aquaticum). Invasive Plant Sci. Manage., 3, 262-267.
Wersal R.M., Madsen J.D., Woolf T.E. & Eckberg N., 2010. Assessment of herbicide efficacy on Eurasia watermilfoil and impacts to the native submersed plant community in Hayden Lake, Idaho, USA. J. Aquat. Plant Manage., 48, 5-11.
Wilton-Jones G., 2005. Control of New Zealand pygmyweed Crassula helmsii by covering with black polythene at The Lodge RSPB Reserve, Bedforshire, England. Conserv. Evidence, 2, 63.
To cite this article
About: Emmanuel Delbart
Univ. Liège - Gembloux Agro-Bio Tech. Unité Biodiversité et Paysage. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
About: Grégory Mahy
Univ. Liège - Gembloux Agro-Bio Tech. Unité Biodiversité et Paysage. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
About: Arnaud Monty
Univ. Liège - Gembloux Agro-Bio Tech. Unité Biodiversité et Paysage. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail : arnaud.monty@ulg.ac.be
