


- Portada
- volume 16 (2012)
- numéro 2
- Définition du profil écologique de l'azobé, Lophira alata, une espèce ligneuse africaine de grande importance : synthèse bibliographique et perspectives pour des recherches futures


Vista(s): 7354 (122 ULiège)
Descargar(s): 421 (8 ULiège)

Définition du profil écologique de l'azobé, Lophira alata, une espèce ligneuse africaine de grande importance : synthèse bibliographique et perspectives pour des recherches futures

Notes de la rédaction
Reçu le 20 juin 2011, accepté le 16 mars 2012
Résumé
L'effort de gestion durable consenti depuis plus de deux décennies dans les forêts denses humides africaines continue d'être confronté à l'insuffisance des connaissances de ces écosystèmes. L'écologie des essences ligneuses demeure peu étudiée, rendant difficile toute possibilité de gestion durable sur le long terme. Cette situation est illustrée à travers la présente synthèse bibliographique sur l'écologie et la sylviculture de l'azobé, Lophira alata Banks ex C.F.Gaertn. (Ochnaceae), une espèce africaine de grande importance jugée « vulnérable » par l'UICN. La revue de littérature proposée met en exergue l'incertitude qui perdure sur la taxonomie de cette espèce, mais aussi de nombreuses insuffisances ou méconnaissances relatives à sa biologie de reproduction, ses conditions de régénération, ses paramètres de la dynamique des populations, ainsi que l'organisation spatiale de sa diversité génétique. Par manque d'informations sur ses exigences écologiques et les facteurs ayant historiquement influencé sa dynamique des populations, les essais sylvicoles menés à ce jour révèlent des résultats hasardeux qui restent pour majeure partie très mitigés. Remédier à ces diverses lacunes par une meilleure caractérisation de son écologie et des essais sylvicoles novateurs constituerait une contribution notable pour la gestion durable de ses populations.
Abstract
Definition of the ecological profile of Lophira alata (ekki), a major important African timber species: literature review and perspectives for future studies. Over two decades, labors made in order to promote sustainable management of African tropical forests are tackled to the lack of knowledge about this complex ecosystem. Ecological parameters of timber species are no more studied, this complicates long-term sustainable forest management. This literature review related to the ecology and silviculture of ekki, Lophira alata Banks ex C.F.Gaertn. (Ochnaceae), a main African timber species recorded as “vulnerable” in the IUCN Red List, will serve as a study’s case. The planned literature review reveals the doubt about its taxonomy, as well as the lack of understanding concerning its reproductive biology, growth conditions, population dynamics parameters, and the spatial distribution of its genetic diversity. The deficiency of knowledge about its ecological needs and the factors which have historically influenced its population dynamics explain why most silvicultural trails provide hazardous and uncertain results. Remedying these gaps in order to improve its ecological characterization and innovative silvicultural trails, would be a significant contribution to the sustainable management of its populations.
Tabla de contenidos
1. Introduction
1Les forêts denses humides tropicales abritent plus de 50 % de la biodiversité terrestre et environ 20 % des espèces végétales (Dupuy, 1998). Face à leur disparition accélérée et aux enjeux qui en découlent, notamment en termes d'émissions de gaz à effet de serre et d'érosion de la biodiversité, celles-ci sont au centre d'importantes préoccupations internationales (FAO, 2009 ; Smith et al., 2011).
2Les forêts denses humides africaines couvrent près de 236 millions d'hectares et régressent à raison de 0,23 % par an (Mayaux et al., 2003 ; FAO, 2010). Cette déforestation s'explique essentiellement par la propagation de l'agriculture permanente à petite échelle (FAO, 2009). En évitant la conversion des forêts en terres agricoles, l'exploitation forestière durable peut être une alternative crédible conciliant préservation du couvert forestier et développement économique des populations locales (Siry et al., 2005). On estime qu'actuellement un effort de gestion durable est consenti sur quelque 30 millions d'hectares, dont 7,6 millions font l'objet d'une certification indépendante (FAO, 2011 ; FSC, 2011). Ces écosystèmes restent toutefois insuffisamment connus (de Wasseige et al., 2009). L'écologie des essences commerciales demeure peu étudiée et des paramètres fondamentaux de leur dynamique des populations restent imprécis (Doucet, 2003), rendant illusoire toute volonté de modélisation sur le long terme. En conséquence, l'information permettant aux gestionnaires forestiers de prendre des décisions adéquates n'est pas toujours disponible (FAO, 2011) et la durabilité tant recherchée en est affectée. L'azobé (ekki en anglais), Lophira alata Banks ex C.F.Gaertn., n'échappe pas à ce constat. En raison de son exploitation commerciale, localement importante, l'UICN la classe dans la catégorie des espèces « vulnérables » (IUCN, 2011). Cet article a pour objectif de proposer une synthèse des connaissances acquises sur l'autoécologie de l'azobé en vue de :
3– mieux caractériser son profil écologique,
4– proposer des perspectives pour des études ultérieures garantissant sa gestion durable.
5Un accent est particulièrement porté sur les aspects d'autoécologie fondamentale et sur les domaines qui, dans une certaine mesure, peuvent faire l'objet d'application à court terme.
2. Considérations taxonomiques et description botanique
2.1. Considérations taxonomiques
6Le genre Lophira de la famille des Ochnaceae est endémique au continent africain. Au début du siècle dernier, Chevalier (1909) distinguait deux espèces : Lophira procera A.Chev., pour l'espèce de forêt et Lophira alata Banks ex C.F.Gaertn., pour l'espèce de savane. Ensuite, Hutchinson et al. (1954) ont montré que L. alata était synonyme de L. procera, et que l'espèce de savane devait se nommer Lophira lanceolata Tiegh. ex Keay.
2.2. L'arbre : présentation botanique
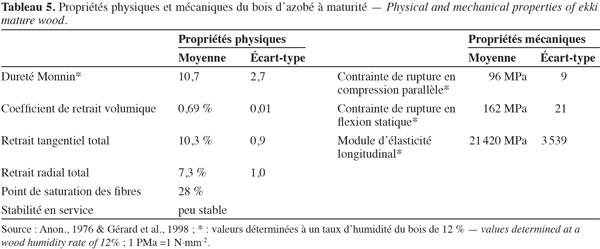
7Les informations qui suivent proviennent de Van Tieghem (1901), Chevalier (1909), Letouzey (1957), Aubréville (1959), de la Mensbruge (1966), Bamps (1967), Anon. (1976), Vivien et al. (1985), White et al. (1996) et Palla et al. (2002). L'azobé peut atteindre 60 m de hauteur et 180 cm de diamètre. Son fût élevé, cylindrique, est parfois un peu sinueux (Figure 1). Sa base est légèrement épaissie et sans contreforts. L'arbre est bas-branchu en lisière. L'écorce de teinte grisâtre à rougeâtre avec une mince pellicule jaune soufre vers l'extérieur atteint 20 mm d'épaisseur. Elle est granuleuse à la cassure et s'exfolie en plaques irrégulières. Sa couche interne, blanchâtre, avec des fibres orangées, est mince et lisse. La cime hémisphérique est fortement charpentée (Figure 1). Les rameaux présentent des cicatrices foliaires très apparentes. Les feuilles, de couleur rouge vif à l'état jeune, sont disposées en rosettes aux extrémités des rameaux. Elles sont alternes, simples, à stipules entières lancéolées-acuminées de 5 mm de long et 2 mm de large. Le pétiole, court et cylindrique, glabre, mesure moins de 2,6 cm de long. Le limbe oblong-obovale, à sommet arrondi ou émarginé, à base cunéiforme, mesure 10 à 25 cm de long et 3,5 à 12 cm de large ; il peut atteindre 50 cm de long, parfois plus, chez les plantules. Il est entier, glabre, avec une nervure médiane saillante sur les deux faces. Les nervures latérales sont nombreuses, rapprochées, parallèles, en relief sur les deux faces et réunies par un réseau apparent de nervilles.

8L'azobé est hermaphrodite. Les panicules terminales, lâches, pyramidales, glabres, mesurent 12 à 20 cm de long. Elles sont à axes anguleux et striés longitudinalement. Les fleurs blanches, odorantes, ont cinq sépales imbriqués persistants, cinq pétales libres et sessiles et de très nombreuses étamines à filets courts de 5 à 7 mm de long. Les pétales suborbiculaires, à onglet large et court, échancrés au sommet, font l7 mm de long et 14 mm de large. Les étamines sont disposées en quatre rangées concentriques. Les anthères oblongues, jaune-orangé, de 4 à 6 mm de long, s'ouvrent par deux ports apicaux. L'ovaire en cône allongé, entier, uniloculaire, se termine par deux stigmates aigus divergents. Sa surface glabre est plissée longitudinalement. Sa loge contient une douzaine d'ovules très courts disposés en deux rangées et insérés sur un axe central. Les deux sépales extérieurs forment dans le fruit deux ailes dissymétriques, rougeâtres à l'état frais, coriaces, nervillées, oblongues et de longueurs inégales : 8 cm pour la petite et 15 cm pour la grande. Le fruit est un akène subligneux et conique de 3 cm de long qui sèche et brunit à maturité. Il contient une seule graine ovoïde et oléagineuse. L'embryon, dépourvu d'endosperme, renferme des cotylédons charnus et épais. Sa germination se produit par apparition d'un épicotyle sortant en crosse. La graine engendre fréquemment deux tigelles, parfois trois.
9L'enracinement de la plantule est pivotant. Les racines latérales sont très fines et nombreuses. Les premières feuilles de la plantule sont opposées et les feuilles suivantes sont disposées sur de courts entrenœuds dans un plan perpendiculaire. Elles sont finement nervurées latéralement. Leur limbe est à marges régulières. L'épicotyle glabre, à feuilles non avortées, mesure entre 13 et 17 cm de long.
2.3. Différenciation taxonomique avec L. lanceolata
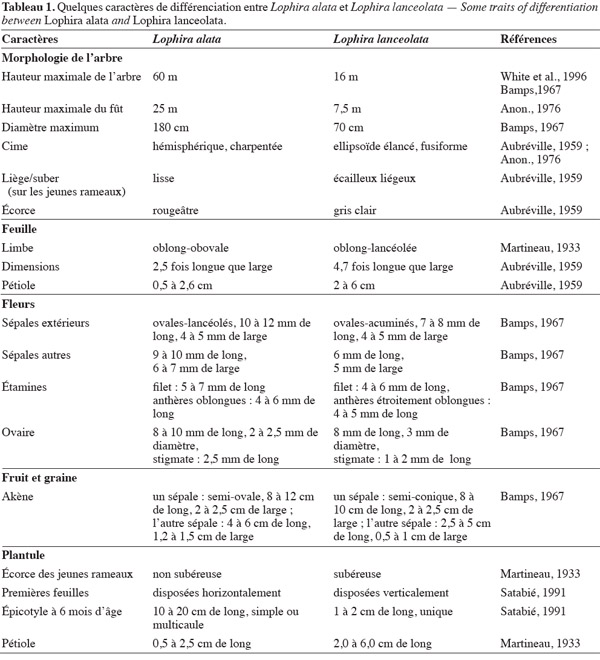
10La distinction des deux espèces du genre Lophira, à savoir L. alata (azobé) et L. lanceolata (méné), a souvent été sujette à controverse (Anon., 1954). Van Tieghem (1901), en examinant la morphologie et les dimensions foliaires des deux taxa de forêt et de savane, ainsi que ceux des formes juvéniles et adultes, avait séparé les Lophira en plusieurs espèces. Hutchinson et al. (1954) mentionnent qu'il s'agit de deux espèces écophylétiques, l'une (méné) commune aux savanes soudano-guinéennes et l'autre (azobé) caractéristique des forêts denses humides guinéo-congolaises. Toutes deux seraient communes dans leurs habitats respectifs et se distingueraient par des particularités morphologiques (Aubréville, 1959). Ces singularités se rapporteraient surtout aux aspects macromorphologiques tels que la conformation des arbres, les dimensions des limbes et pétioles, les dimensions des sépales et stigmates, la largeur des sépales des fruits et la morphologie des plantules (Tableau 1). Toutefois, ces éléments resteraient très insuffisants pour en faire deux espèces distinctes (Aubréville, 1959). Les formes juvéniles de ces deux taxa seraient difficiles à distinguer en lisière de forêt dense humide où L. alata et L. lanceolata se côtoient (Letouzey, 1985). Leur génotype serait également identique, leur pollen et leur nombre chromosomique (2n égal à 28) paraissent identiques (Satabié, 1991).
3. Distribution et origine des peuplements forestiers de L. alata
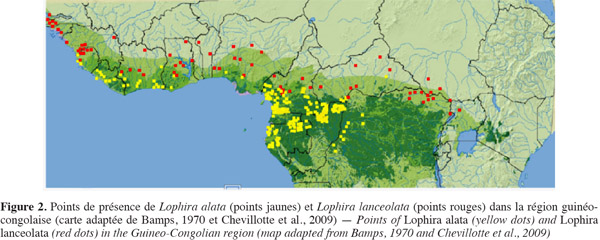
11L'azobé a une distribution omni-guinéo-congolaise (Doucet, 2003). Son aire de répartition s'étend du Congo depuis le district forestier central jusqu'en Sierra Leone voire en Guinée Bissau et ne descend pas en dessous du bassin inférieur de l'Ogooué au Gabon (Bamps, 1967 ; Anon., 1976). Il se rencontre au contact de la savane, dans les forêts denses sempervirentes, sur les pentes de montagne et dans les terrains marécageux (Aubréville, 1959 ; Bamps, 1970 ; White et al., 1996). C'est une espèce des forêts secondaires « ombrophiles semi-sempervirentes humides mélangées » et « ombrophiles sempervirentes littorales hygrophiles » (White, 1983). C'est au sein de cette dernière qu'elle présenterait les densités les plus élevées, soit plus de deux arbres (dbh supérieur ou égal à 40 cm) à l'hectare (Guillaumet et al., 2010). En Afrique centrale atlantique, la distribution de l'azobé serait surtout limitée aux forêts denses humides, l'espèce ne se rencontrant dans les forêts secondaires qu'au voisinage des grands cours d'eau (Figure 2).
12Plusieurs hypothèses ont été évoquées pour expliquer l'origine des populations de L. alata en forêt dense humide. Letouzey (1968) avait suggéré une origine paléobotanique et anthropique fondée sur les nombreuses ressemblances relevées avec L. lanceolata et le rapprochement des aires de répartitions de ces deux espèces. Selon cet auteur, L. alata résulterait d'une adaptation écologique de L. lanceolata en forêt dense humide, puis se serait propagé dans cette région à la suite des défrichements humains.
13Maley (1994), sans valider cet argument anthropique, met en exergue l'impact des variations paléo-climatiques sur la dynamique des populations d'azobé. Après la régression majeure des forêts sous l'effet d'un climat aride entre 20 000 et 14 000 ans BP (before present), le retour à un climat plus humide de 14 000 à 9 500 ans BP a été marqué par la recolonisation des méso- et méga-phanérophytes typiques des formations forestières guinéo-congolaises, dont les populations d'azobé (Maley, 1994 ; Maley et al., 1997). Vers 3 000 ans BP, les régions en bordure du golfe de Guinée auraient connu un assèchement qui a provoqué de vastes ouvertures et un recul de la forêt (Maley, 1994). Ces ouvertures auraient permis la migration vers le Sud des populations Bantou (Schwartz, 1992). En combinaison avec les actions anthropiques, notamment les défrichements et les incendies à des fins agricoles (Schwartz et al., 1995), cet assèchement climatique aurait renforcé l'installation d'une végétation de type mosaïque forêt-savane (Elenga et al., 1992) et l'extension des taxons pionniers (Maley, 2001). Le rôle crucial de l'homme dans la dynamique des populations d'azobé a été souligné par Letouzey (1985) qui a établi un rapprochement entre l'abondance de fragments de poteries et de charbons de bois au pied de spécimens de L. alata en forêt littorale camerounaise et la présence de l'espèce dans cette région. L'extension typique de la forêt climacique n'a repris que vers 2 000 ans BP (Maley, 1994).
4. Exigences climatiques, topographiques et pédologiques
14L'aire de l'azobé, limitée à la région guinéo-congolaise, s'étend aux climats tropicaux de types humide (Af) et de mousson (Am). Les précipitations annuelles moyennes y sont comprises entre 1 600 et 2 000 mm, excepté dans les zones côtières de la haute et basse Guinée et une partie du bassin du Congo (RDC) qui en reçoivent davantage (White, 1983). Les températures moyennes mensuelles varient entre 24 et 27 °C (White, 1983). L'azobé abonde au bord des côtes atlantiques (Letouzey, 1957). Il y forme des peuplements étendus et pénètre à l'intérieur du continent dans les vallées et le long des grands cours d'eau (Anon., 1954). Il ne s'élève guère à plus de 800 m d'altitude sur les pentes des montagnes (Anon., 1976 ; Vivien et al., 1985). Sa distribution semble dépendre surtout des conditions climatiques et édaphiques qui règnent dans son aire de répartition (Veenendaal et al., 1998). En effet, selon ces auteurs, la saison sèche constituerait un important facteur de limitation de sa répartition. L'azobé manifesterait une préférence envers les sols à hydromophie de profondeur et présenterait une meilleure régénération sur des sols humides bien drainés (Taylor, 1960 ; Dupuy, 1998). C'est une espèce rustique qui tend à préférer les sols sablonneux et sablo-argileux à nappe phréatique peu profonde (Anon., 1954 ; Letouzey, 1957 ; Aubréville, 1959 ; Letouzey, 1979). L'espèce préfère les sols pauvres et acides (Swaine et al., 19941 cités par Hawthorne, 1995) où elle forme des associations mycorhiziennes (Onguene et al., 2001). Les semis présentent des meilleurs taux de croissance dans les sols peu fertiles, en raison probablement de leur incapacité à profiter de la disponibilité en éléments nutritifs (Baker et al., 2003). L'espèce constituerait d'ailleurs un bon indicateur de sols pauvres au sud du Cameroun (Kanmegne, 2004).
5. Caractéristiques écologiques
5.1. Tempérament et structure des populations
15Selon divers auteurs (Aubréville, 1959 ; Hawthorne, 1995 ; Doucet, 2003), le tempérament de l'azobé est qualifié d'héliophile modéré à pionnier. Un ombrage latéral serait préférable les premiers mois, mais les plantules auraient rapidement besoin d'un éclairement relatif de 50 à 90 % (Catinot, 1965). En effet, les semis d'azobé présenteraient des taux de mortalité élevés induits par la sècheresse, soit plus de 80 % pendant la phase d'établissement (Baker et al., 2003). Lorsque les plantules sont maintenues de façon prolongée dans un environnement ombragé, leurs taux de croissance paraissent diminuer (Voorhoeve, 1965). Les juvéniles, contrairement aux semis et aux plantules, présenteraient un meilleur développement en bordure des trouées d'abattage au Sud du Cameroun (Bongjoh et al., 2001). Ainsi, ces exigences se traduiraient par des déficits locaux de régénération (Letouzey, 1957), bien que la situation puisse être fort variable d'une forêt à une autre, probablement en raison de passés anthropiques différents (importances des milieux agricoles). Les structures diamétriques qui en résultent évoluent d'une courbe de Gauss à une exponentielle décroissante (Sépulchre et al., 2008).
5.2. Phénologie
16En se référant aux observations d'Anon. (1954), Taylor (1960), Voorhoeve (1965), Hecketsweiler (1992), White et al. (1996), Yalibanda (1999), Doucet (2003) et Eyog Matig et al. (2006), la phénologie de l'azobé parait présenter un assez bon conservatisme sur toute son aire de répartition, bien que ses phénophases pourraient correspondre à des climats différents. En effet, dans les régions soumises à un climat tropical humide (Af), notamment au Cameroun et en République Centrafricaine, l'azobé se défeuille durant une à deux semaines entre novembre et décembre au début de la saison sèche. La floraison qui s'ensuit se produit de manière régulière et abondante entre décembre et janvier en saison sèche. La fructification s'étale sur trois à quatre mois dès janvier voire février pendant la saison sèche jusqu'en avril ou mai, au début de la saison des pluies. Sous les climats tropicaux de mousson (Am), notamment au Liberia jusqu'au Gabon ainsi qu'en RDC, l'espèce est à feuilles caduques entre octobre et décembre pendant la grande saison des pluies. Les arbres fleurissent entre novembre et janvier dès la fin de la grande saison des pluies, durant la grande saison sèche. Une exception est à relever au Gabon où cette floraison se fait durant la grande saison des pluies, entre décembre et janvier. Le développement des fruits se fait entre janvier et mars voire avril, dès la fin de la grande saison sèche, pendant la petite saison des pluies. Les fruits matures apparaissent entre mars et juin, pendant la petite saison des pluies. Au Gabon, celle-ci se fait entre février et mars pendant la petite saison sèche.
17Selon Yalibanda (1999), les arbres fleurissent en forêt dense humide semi-sempervirente en République Centrafricaine dès 15 cm de diamètre. Cette floraison devient régulière en forêt littorale camerounaise à partir de 50 cm de diamètre (Anon., 1954). Doucet (2003) observe en forêt sempervirente côtière hygrophile au Gabon que les arbres fructifient dès 34 cm de diamètre et produisent régulièrement des fruits dès 45 cm de diamètre.
5.3. Régénération naturelle
18L'azobé est une espèce ptérochore avec une dispersion anémochore des fruits (Doucet, 2003). Dans les conditions favorables (par grand vent), ceux-ci peuvent être transportés jusqu'à 200 m du semencier (Anon., 1954). En conséquence, les semis s'installent préférentiellement dans la direction des vents dominants. C'est une espèce sociale qui se régénère aisément dans les endroits découverts. Toutefois, cette régénération semble exiger une absence totale d'espèces concurrentes (Letouzey, 1957). L'espèce rejette bien des souches mais ne drageonne pas (Anon., 1954). Les fruits seraient consommés par les ruminants et les rongeurs, ces derniers déterrant et consommant les cotylédons (Anon., 1954). Les bourgeons des plantules seraient détruits par un coléoptère, Tragocephala guerenii White (Foahom, 2002), tandis que les feuilles seraient consommées par diverses chenilles dont Imbrasia obscura Butler et Antheua insignata Gaede (Foahom, 2002).
5.4. Accroissements annuels moyens en diamètre et en hauteur
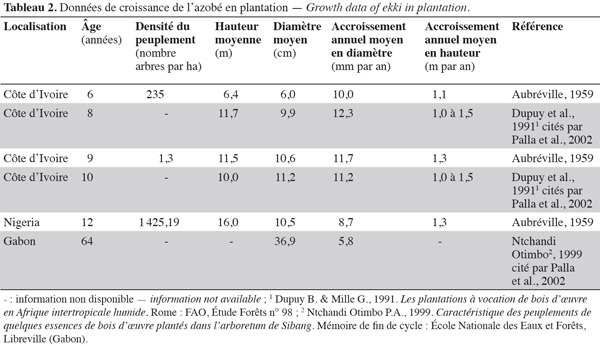
19Les données relatives à l'accroissement annuel moyen en diamètre demeurent très lacunaires. En forêt naturelle, cet accroissement varie de 4 à 8 mm par an pour la Sierra Leone, le Ghana, le Nigeria, le Congo et le Gabon (Savill et al., 19672 cités par Hawthorne, 1995 ; Dupuy, 1998 ; Doucet, 2003). En plantation, cet accroissement se situe entre 6 et 12 mm par an (Tableau 2). Aubréville (1959) mentionne des accroissements annuels moyens en diamètre pouvant atteindre 15 mm par an chez les jeunes arbres exposés aux conditions optimales d'éclairement. L'accroissement annuel moyen en hauteur est généralement compris entre 1,0 et 1,5 m par an (Tableau 2). Ces informations restent toutefois absentes dans d'autres régions, notamment au Cameroun. Il en est de même de la nature et de la périodicité des cernes de croissance qui, jusqu'à présent, n'ont pas été établies.
6. Sylviculture et aménagement forestier
6.1. Législations forestières
20Deux mesures de l'arsenal légal des pays tropicaux concernent directement ou indirectement l'azobé (Sépulchre et al., 2008). La première consiste en la fixation, dans chaque pays producteur, de diamètres minima d'exploitation (DME). Ils correspondent au seuil légal en dessous duquel aucun pied ne peut être exploité (Tableau 3). La seconde repose sur les taux de reconstitution minima à atteindre. Au Cameroun, ce taux ne doit pas être inférieur à 50 %.

6.2. Tarifs de cubage
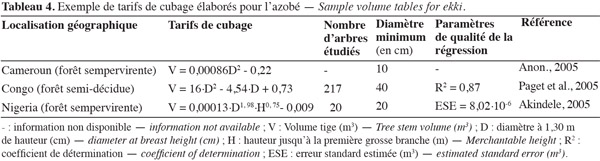
21Pour des raisons commerciales ou d'études de productivité forestière, différents tarifs de cubage ont été développés pour l'azobé (Tableau 4). Ceux-ci permettent d'estimer le volume brut d'une tige sur la base de son diamètre et/ou de sa hauteur. Leur validité se limite à la zone couverte par l'échantillon d'arbres étudiés et au diamètre minimum de prise en compte des arbres (Paget et al., 2005). Le fait que la taille des échantillons d'arbres étudiés soit limitée à quelques individus (Akindele, 2005), que les zones couvertes par ces échantillons s'étalent sur plusieurs millions d'hectares (Paget et al., 2005) et que certains auteurs ne donnent pas d'information sur la méthode de construction de leur modèle (Anon., 2005), amène à être prudent quant à l'utilisation de ces formules.
6.3. Plantations et régénération artificielle
22L'azobé a un taux de germination élevé entre 85 et 95 % (de la Mensbruge, 1966). Les graines, dont la masse moyenne est d'environ 0,3 g, ne se conservent pas, bien que quatre mois après la récolte, le taux de germination soit encore d'environ 25 % (Anon., 1954 ; Taylor, 1960). La germination, hypogée et échelonnée, dure en moyenne 18 à 30 jours pour les graines à embryon unique et peut atteindre 45 jours dans le cas de polyembryonie (de la Mensbruge, 1966). Lors du semis en pépinière, les graines doivent être couchées et enterrées sous 1 cm de substrat pour faciliter la sortie de la tigelle (Anon., 1954). Les plants sont utilisés, en saison des pluies, après 15 à 18 mois d'élevage, lorsqu'ils ont une hauteur de 40 à 50 cm et 15 feuilles environ (Anon., 1954 ; Taylor, 1960).
23Les rares essais sylvicoles réalisés font apparaitre des résultats mitigés. Dès 1930, en Côte d'Ivoire, l'azobé a été planté en forêt naturelle dans des layons étroits à grand écartement en vue d'obtenir une densité de 50 arbres à l'hectare (Dupuy, 1992). Cinquante années après, les densités relevées varient entre 6 et 28 tiges·ha-1 (Dupuy, 1992). En 1935 au Gabon, une parcelle d'azobé a été plantée à raison de 1 100 tiges·ha-1, et 64 ans après, seules 128 tiges·ha-1 subsistaient (Palla et al., 2002).
7. Caractéristiques, utilisations et commerce international du bois
7.1. Caractéristiques du bois et utilisations industrielles
24Les informations qui suivent sont de Méniaud (1950), Anon. (1954), Anon. (1976) et Gérard et al. (1998). Le bois d'azobé présente d'excellentes propriétés physiques et mécaniques (Tableau 5). Il a une bonne résistance vis-à-vis des champignons de pourriture. Le bois parfait est résistant aux attaques d'insectes. La durabilité du bois vis-à-vis des termites est bonne. Il est considéré comme un bois non imprégnable (classe d'imprégnabilité : 4) selon la norme NF EN 350-2 (norme européenne sur la durabilité du bois et des produits dérivés du bois). Le bois d'azobé est apprécié pour les installations en milieu exposé tels les ouvrages portuaires, les constructions hydrauliques, les traverses de chemin de fer et les fonds de wagon. Il est employé pour la fabrication de seuils, pièces d'appui, jets d'eau et pour certains éléments de maisons à ossature en bois. Il convient à la fabrication de parquets et escaliers destinés à des applications industrielles.
7.2. Usages traditionnels
25Les informations qui suivent sont d'Anon. (1954), Bouquet (1969), Kanmegne (2004), Kayode (2006), Eyog Matig et al. (2006) et Ibrahim et al. (2007). Bien que des confusions dans les usages soient récurrentes avec L. lanceolata, l'azobé parait ne pas fournir de produits accessoires dont l'utilisation serait généralisée. En effet, l'huile extraite des graines serait la seule partie de l'arbre qui interviendrait dans l'alimentation au Cameroun. Le bois d'azobé serait utilisé localement dans ce pays pour la confection des ponts. Les chenilles comestibles Imbrasia obscura (Butler, 1878) dont l'arbre hôte est Piptadenia africana Hook.f. (Dabema), seraient présentes sur l'azobé en RDC. La pharmacopée traditionnelle, qui reconnait de nombreuses vertus à certaines parties de l'arbre, serait l'utilisation la plus répandue, notamment au Gabon, au Nigeria, en RCA et en RDC. L'écorce est employée dans le traitement de diverses maladies (caries dentaires, douleurs viscérales, fièvre jaune, maux de cœur, etc.). Récoltée sur le tronc, elle constituerait un analgésique et un anti-inflammatoire. Mélangée à l'écorce d'Afzelia africana Sm. ex Pers., elle servirait à lutter contre les troubles mentaux. Les feuilles, les racines et les graines permettraient de lutter contre le paludisme. Les jeunes feuilles seraient utilisées contre les troubles respiratoires. Les feuilles adultes, en mulch, permettraient de lutter contre les termites. Ainsi, l'azobé, qui parait ne pas présenter une grande valeur économique en tant que produit non ligneux, serait surtout recherché au niveau traditionnel pour ses vertus médicinales.
7.3. Commerce international du bois
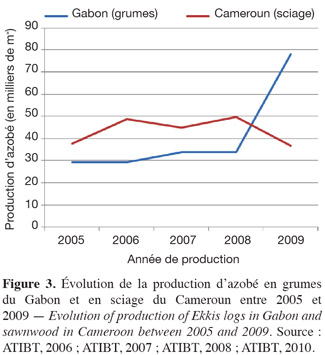
26L'azobé produit un bois de grande valeur commerciale. Entre 2005 et 2009, ses exportations ont surtout été réalisées par le Cameroun, le Congo, la Côte d'Ivoire et le Gabon (ATIBT, 2010). Durant cette période, le plus grand exportateur de grumes était le Gabon (en moyenne 41 000 m3 par an) et le plus grand exportateur de sciage était le Cameroun (en moyenne 44 000 m3 par an). Dans ce dernier pays, l'évolution de la production de sciage au cours de ces cinq dernières années s'est maintenue autour de 45 000 m3, bien qu'elle ait affiché une baisse en 2009, sans doute due à la crise financière de 2008 (Figure 3). Au Gabon, la production de grumes, qui variait faiblement autour de 30 000 m3, est passée au-delà du double en 2009 en raison de l'augmentation de la demande asiatique (Figure 3). Les importations du bois d'azobé ont été dominées en 2009 par les Pays-Bas (soit 84,5 % du sciage) et la Belgique (44 % des grumes). Celles-ci devraient logiquement s'accroître à court et moyen termes suite à la demande asiatique qui affiche une tendance à la hausse (ATIBT, 2010).
8. Conclusion et orientations pour des recherches futures
27La présente synthèse bibliographique contribue à dresser le bilan des connaissances existantes sur l'écologie et la sylviculture de l'espèce Lophira alata. Il s'agit d'une analyse critique qui vise à mettre en exergue les domaines d'études suffisamment bien documentés, et ceux qui nécessitent des efforts de recherches futures. Notre analyse fait ressortir un certain nombre d'aspects relativement bien documentés :
28– la caractérisation botanique de l'espèce parait suffisante et fournit des éléments qui permettent de la différencier des taxons de la même famille ;
29– la phénologie et les taux de croissance donnent des tendances générales sur la dynamique reproductive et populationnelle de l'espèce.
30Toutefois, de nombreuses hypothèses restent à vérifier, notamment en ce qui concerne l'origine des populations d'azobé en milieu de forêt dense humide tropicale. Selon Letouzey (1968) et Satabié (1991), L. alata serait un écotype issu d'une adaptation écologique de L. lanceolata, espèce de savane botaniquement et génétiquement proche de L. alata. Si cette hypothèse était vérifiée, elle supposerait que des flux de gènes soient toujours possibles entre ces deux taxa. La caractérisation de l'importance de leurs relations phylogénétiques répondrait non seulement à une question fondamentale sur l'origine de certaines espèces de forêts tropicales humides, mais servirait aussi à reconsidérer la disponibilité et la distribution du genre Lophira en Afrique. Par ailleurs, l'azobé se serait répandu dans le massif forestier africain à la faveur des variations paléoclimatiques et des perturbations anthropiques passées, ces dernières ayant pu avoir un impact particulièrement prépondérant. Mais cela n'a encore jamais été rigoureusement confirmé. Un certain nombre d'autres informations clés sur ses caractéristiques écologiques et génétiques demeurent inconnues, alors qu'elles participeraient certainement à une meilleure gestion de l'espèce sur le long terme :
31– les diamètres minimum et optimal de fructification,
32– les facteurs climatiques initiant la floraison (sensibilité de l'espèce au changement climatique),
33– la cartographie de la diversité génétique de ses populations, permettant de détecter entre autres des populations à risque.
34Dans les zones où l'espèce affiche un déficit de régénération du fait de son tempérament héliophile, un des premiers points à éclaircir est sans doute la quantification de ses besoins réels en lumière. À l'instar d'autres espèces considérées jusqu'alors comme héliophiles strictes, il n'est pas impossible que l'azobé supporte en réalité une certaine pénombre durant les premières années de son développement, chose qui pourrait favoriser en définitive son établissement dans des chablis forestiers de dimensions moyennes. Seule une caractérisation subséquente de la dynamique de sa régénération couplée à des tests sylvicoles permettront de définir les conditions abiotiques optimales pour son développement. Enfin, toujours dans un souci de gestion durable des peuplements d'azobé, les aménagistes forestiers manquent d'informations pragmatiques indispensables pour une modélisation de la dynamique de ses populations à long terme : taux de croissance dans différentes classes de diamètre, taux de mortalité et taux de recrutement des arbres d'avenir sont autant de paramètres à déterminer.
35Remerciements
36Les auteurs remercient l'Université de Douala (Cameroun), l'Université de Liège (Belgique), l'ERAIFT (Kinshasa) et l'ASBL Nature+ (Belgique) pour les soutiens technique, scientifique et financier accordés aux travaux de doctorat d'Achille Biwole. Leurs remerciements s'adressent également à la société WIJMA Cameroun S.A., structure partenaire, pour son appui aux travaux de terrain. Enfin, que Cédric Vermeulen, Jean-François Gillet, Michèle Federspiel et Jean-Yves de Vleeschouwer soient remerciés pour leurs diverses contributions et leurs conseils.
Bibliographie
Akindele O.S., 2005. Volume functions for common timber species of Nigeria's tropical rain forests. Technical Document. Yokohoma, Japan: International Tropical Timber Organisation.
Anon., 1954. Monographie de Azobé, Lophira procera A.Chev. Nogent-sur-Marne, France : Centre Technique Forestier Tropical.
Anon., 1976. Azobé. Bois Forêts Trop., 170, 35-50.
Anon., 2005. Évaluation des ressources forestières nationales du Cameroun, 2003-2004. Rapport. Yaoundé : Ministère des Forêts et de la Faune du Cameroun ; Rome : FAO.
ATIBT (Association Technique Internationale des Bois Tropicaux), 2006. La lettre de l'ATIBT n°24 : statistiques 2005. Paris : Association Technique Internationale des Bois Tropicaux.
ATIBT (Association Technique Internationale des Bois Tropicaux), 2007. La lettre de l'ATIBT n°26 : statistiques 2006. Paris : Association Technique Internationale des Bois Tropicaux.
ATIBT (Association Technique Internationale des Bois Tropicaux), 2008. La lettre de l'ATIBT n°28 : statistiques 2007. Paris : Association Technique Internationale des Bois Tropicaux.
ATIBT (Association Technique Internationale des Bois Tropicaux), 2010. La lettre de l'ATIBT n°32 : statistiques 2009. Paris : Association Technique Internationale des Bois Tropicaux.
Aubréville A., 1959. La flore forestière de la Côte d'Ivoire. 2e éd. Nogent-sur-Marne, France : Centre Technique Forestier Tropical.
Baker T.R., Swaine M.D. & Burslem D.F.R.P., 2003. Variation in tropical forest growth rates: combined effects of functional group composition and resource availability. Perspect. Plants Ecol. Evol. Syst., 6(1-2), 21-36.
Bamps P., 1967. Flore du Congo, du Rwanda et du Burundi : spermatophytes, Ochnaceae. Bruxelles : Jardin Botanique National de Belgique, 54-56.
Bamps P., 1970. Répartition géographique du genre Lophira Banks ex Gaertn. (Ochnaceae). Bull. Jard. Bot. Natl Belg., 40(4), 291-294.
Bongjoh C.A. & Nsangou M., 2001. Gap disturbance regimes and regeneration dynamics of commercial timber tree species in a Southern Cameroon forest. In: Jonkers W.B.J., Foahom B. & Schmidt P., eds. Seminar Proceedings “Sustainable Forest Management of African Rain Forest”, Part II. Symposium, November 1999, Kribi, Cameroon. Wageningen, The Netherlands: The Tropenbos Foundation, 112-124.
Bouquet A., 1969. Féticheurs et médecines traditionnelles du Congo (Brazzaville). Paris : ORSTOM.
Catinot R., 1965. Sylviculture tropicale en forêt dense africaine. Bois Forêts Trop., 103, 3-16.
Chevalier A., 1909. Les végétaux utiles de l'Afrique tropicale française. Première étude sur les bois de la Côte d'Ivoire. Vol. V. Paris : Challamel.
Chevillotte H. et al., 2009. Les essences forestières commercialisées de l’Afrique tropicale humide, http://phyto-afri.ird.fr, (10/06/2011).
de la Mensbruge G., 1966. La germination et les plantules des essences arborées de la forêt dense humide de la Côte d'Ivoire. Nogent-sur-Marne, France : Centre Technique Forestier Tropical.
de Wasseige C. et al., 2009. Les forêts du bassin du Congo. État des forêts 2008. Luxembourg : Office des Publications de l'Union Européenne.
Doucet J.-L., 2003. L'alliance délicate de la gestion forestière et de la biodiversité dans les forêts du centre du Gabon. Thèse de doctorat : Faculté universitaire des Sciences agronomiques de Gembloux (Belgique).
Dupuy B., 1992. Les plantations à vocation de bois d'œuvre en forêt dense humide africaine. Bois Forêts Trop., 213, 5-15.
Dupuy B., 1998. Base pour une sylviculture en forêt dense humide africaine. Série FORAFRI, Document 4. Montpellier, France : CIRAD-Forêt.
Elenga H. et al., 1992. Changements climatiques et action anthropique sur le littoral congolais au cours de l'Holocène. Bull. Soc. Géol. Fr., 163(1), 83-90.
Eyog Matig O., Ndoye O., Kengue J. & Awono A., 2006. Les fruitiers forestiers comestibles du Cameroun. Cotonou : International Plant Genetic Resources Institute.
FAO, 2009. Situation des forêts du monde 2009. Rome : FAO.
FAO, 2010. Évaluation des ressources forestières mondiales 2010. Résultats principaux. Rome : FAO.
FAO, 2011. La situation des forêts dans le bassin amazonien, le bassin du Congo et l'Asie du Sud-Est. Rapport préparé pour le sommet des trois bassins forestiers tropicaux du 31 mai au 3 juin 2011 à Brazzaville, République du Congo. Rome : FAO, OIBT.
Foahom B., 2002. Insect pest incidence on timber tree species in natural forest in South Cameroon. Kribi, Cameroon : The Tropenbos-Cameroon Program.
FSC, 2011. Global FSC certificates: type and distribution. Bonn, Germany : FSC International Center GmbH.
Gérard J. et al., 1998. Synthèse sur les caractéristiques technologiques de référence des principaux bois commerciaux africains. Série FORAFRI, Document 11. Montpellier, France : CIRAD-Forêt.
Guillaumet J.-L. et al., 2010. Atlas des essences commercialisées d'Afrique tropicale humide : l'exemple du Cameroun. In : van der Burgt X., van der Maesen J. & Onana J.-M., eds. Systematics and Conservation of African Plants. Proceedings of the 18th AETFAT Congress, Yaounde, Cameroon. Richmond, UK: Kew Royal Botanic Gardens, 759-770.
Hawthorne W.D., 1995. Ecological profiles of Ghanaian forest trees. Oxford, UK: Oxford Forestry Institute.
Hecketsweiler P., 1992. Phénologie et saisonnalité en forêt gabonaise. L'exemple de quelques espèces ligneuses. Thèse de doctorat : Université Montpellier 2 (France).
Hutchinson J. & Dalziel J.M., 1954. Flora of West Tropical Africa. 2nd ed. London: Crown Agents.
Ibrahim J.A. et al., 2007. Ethno-medicinal plants and methods used by Gwandara tribe of Sabo Wuse in Niger state, Nigeria, to treat mental illness. Afr. J. Trad. Complementary Altern. Med., 4(2), 211-218.
IUCN (International Union for Conservation of Nature and Natural Resources), 2011. IUCN Red list of threatened species, www.iucnredlist.org., (16/06/2011).
Kanmegne J., 2004. Slash and burn agriculture in the humid forest zone of Southern Cameroon: soil quality dynamics, improved fallow management and farmers' perceptions. PhD thesis: Wageningen University and Research Centre (The Netherlands).
Kayode J., 2006. Conservation of indigenous medical botanicals in Ekiti state, Nigeria. J. Zhejiang Univ. Sci. B., 7(9), 713-718.
Letouzey R., 1957. La forêt à Lophira alata de la zone littorale camerounaise. Bois Forêts Trop., 53, 9-20.
Letouzey R., 1968. Étude phytogéographique du Cameroun. Paris : P. Lechevalier.
Letouzey R., 1979. Végétation. In : Laclavère G. et al., eds. Atlas de la République du Cameroun. Paris : Groupe J.A., 20.
Letouzey R., 1985. Notice de la carte phytogéographique du Cameroun au 1 : 500 000. Paris : P. Lechevalier.
Maley J., 1994. Le cadre paléoenvironnemental des refuges forestiers africains : quelques données et hypothèses. In : van der Maesen L.J.G., van der Burgt X.M. & van Medenbach de Rooy J.M., eds. The biodiversity of African plants: proceedings, 14th AETFAT Congress, 22-27 August 1994, Wageningen, The Netherlands. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Maley J., 2001. La destruction catastrophique des forêts d'Afrique centrale survenue il y a environ 2 500 ans exerce encore une influence majeure sur la répartition actuelle des formations végétales. Syst. Geogr. Plant, 71, 777-796.
Maley J. & Brenac P., 1997. Vegetation dynamics, paleoenvironments and climatic changes in the forests of Western Cameroon during the last 28,000 years B.P. Rev. Palaeobot. Palynol., 99, 157-187.
Martineau M., 1933. Notes sur les Lophira de forêt et de savane. Rev. Bot. Appl., 13, 467-469.
Mayaux P. et al., 2003. A land cover map of Africa. Carte de l'occupation du sol de l'Afrique. Luxembourg : European Commission.
Méniaud J., 1950. L'Azobé et ses utilisations. Bois Forêts Trop., 15, 261-266.
Onguene N.A. & Kuyper T.W., 2001. Mycorrhizal associations in the rain forest of South Cameroon. For. Ecol. Manage., 140, 277-287.
Paget D. & Desmet O., 2005. Étude dendrométrique pour l'aménagement des UFA Pokola, Kabo, Loundoungou et Toukoulaka (Nord Congo) : tarifs de cubage, coefficient d'exploitation et de commercialisation, accroissement diamétrique des essences. Document final. Congolaise Industrielle des Bois (TIB) & Tropical Wood Environment (TWE).
Palla F., Louppe D. & Doumenge C., 2002. Azobé. Fiche technique, écologique et sylvicole. Montpellier, France : CIRAD-Forêt.
Satabié B., 1991. Compte-rendu de l'étude de quelques éléments de la biosystématique à l'interprétation de la vicariance des deux espèces de Lophira (Ochnaceae) au Cameroun. Candollea, 46, 85-94.
Schwartz D., 1992. Assèchement climatique vers 3 000 B.P. et extension Bantu en Afrique centrale atlantique : quelques réflexions. Bull. Soc. Géol. Fr., 163(3), 353-361.
Schwartz D. et al., 1995. Les savanes du Congo : une végétation spécifique de l'Holocène supérieur. In : 2e Symposium de Palynologie africaine, 1995, Tervuren, Belgique. Orléans, France : CIFEG, 99-108.
Sépulchre F., Daïnou K. & Doucet J.-L., 2008. Étude de la vulnérabilité de 18 espèces ligneuses commerciales d'Afrique centrale reprises sur la liste rouge IUCN. Rapport d'étude. Gembloux, Belgique : Faculté universitaire des Sciences agronomiques de Gembloux, ASBL Nature+, ATIBT.
Siry P.J., Cubbage W.F. & Miyan Rukunuddin A., 2005. Sustainability forest management: global trends and opportunities. For. Policy Econ., 7, 551-561.
Smith J.M. et al., 2011. Assessing the impacts of international trade on CITES-listed species: current practices and opportunities for scientific research. Biol. Conserv., 144, 82-91.
Taylor C.J., 1960. Synecology and silviculture in Ghana. London: Thomas Nelson and Sons Ltd.
Van Tieghem P., 1901. Sur le genre Lophire considéré comme type d'une famille distincte : les Lophiracées. J. Bot., 15, 168-194.
Veenendaal E.M. & Swaine M.D., 1998. Limits to tree species distributions in lowland tropical rainforest. In: Newbery D.M., Prins H.H.T. & Brown M., eds. Dynamics of tropical communities. Oxford, UK: Blackwell Sciences, 163-191.
Vivien J. & Faure J.J., 1985. Arbres des forêts denses d'Afrique centrale. Paris : Agence de Coopération Culturelle et Technique.
Voorhoeve A.G., 1965. Liberian high forest trees. Wageningen, The Netherlands: Centre for Agricultural Publication and Documentation.
White F., 1983. The vegetation of Africa: a descriptive memoir to accompany the UNESCO/AETFAT/UNSO vegetation map of Africa. Paris : UNESCO.
White L. & Abernethy K., 1996. Guide de la végétation de la Réserve de la Lopé. Libreville : ECOFAC Gabon.
Yalibanda Y., 1999. Phénologie en forêt dense de Ngotto (RCA) : bilan de trois années d'observation. In : Actes du séminaire Forafri de Libreville, Gabon. CD-Rom « La gestion des forêts denses africaines aujourd'hui ». Montpellier, France : CIRAD-Forêt.
Notes
Para citar este artículo
Acerca de: Achille Bernard Biwolé
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Gestion des Ressources forestières et des Milieux naturels. Laboratoire de Foresterie des Régions tropicales et subtropicales. Passage des Déportés, 2. B-5030 Gembloux (Belgique). E-mail : achille.biwole@doct.ulg.ac.be – Université de Douala. École normale supérieure d'Enseignement technique. B.P. 1872. CAM-Douala (Cameroun) – École Régionale Post-Universitaire d’Aménagement et de Gestion Intégrés des Forêts et Territoires Tropicaux (ERAIFT). B.P. 15373 Kinshasa (RDC).
Acerca de: Nils Bourland
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Gestion des Ressources forestières et des Milieux naturels. Laboratoire de Foresterie des Régions tropicales et subtropicales. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Kasso Daïnou
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Gestion des Ressources forestières et des Milieux naturels. Laboratoire de Foresterie des Régions tropicales et subtropicales. Passage des Déportés, 2. B-5030 Gembloux (Belgique).
Acerca de: Jean-Louis Doucet
Univ. Liège - Gembloux Agro-Bio Tech. Unité de Gestion des Ressources forestières et des Milieux naturels. Laboratoire de Foresterie des Régions tropicales et subtropicales. Passage des Déportés, 2. B-5030 Gembloux (Belgique) – École Régionale Post-Universitaire d’Aménagement et de Gestion Intégrés des Forêts et Territoires Tropicaux (ERAIFT). B.P. 15373 Kinshasa (RDC).
