


- Accueil
- volume 13 (2009)
- numéro 4
- Effets de la fertilisation azotée, de l’inoculation par Rhizobium sp. et du régime des pluies sur la production de la biomasse et la teneur en azote du pois chiche


Visualisation(s): 12081 (53 ULiège)
Téléchargement(s): 662 (6 ULiège)

Effets de la fertilisation azotée, de l’inoculation par Rhizobium sp. et du régime des pluies sur la production de la biomasse et la teneur en azote du pois chiche

Notes de la rédaction
Reçu le 26 novembre 2008, accepté le 12 juin 2009
Résumé
La production du pois chiche dans les régions méditerranéennes est largement dépendante de la disponibilité en eau et en azote. Dans ces régions, le climat est caractérisé par une faible pluviométrie souvent mal répartie et une minéralisation rapide de la matière organique. Afin d’étudier l’effet de la fertilisation azotée et de l’inoculation par des bactéries symbiotiques sur la production de la biomasse et la quantité d’azote accumulée du pois chiche, des essais ont été menés au champ dans deux stations de l’INRAT au nord de la Tunisie durant deux campagnes agricoles, 2000-2001 et 2001-2002, caractérisées par des pluviométries contrastées. Les résultats ont montré que la fertilisation azotée a entrainé une augmentation de la biomasse aérienne et une diminution de la biomasse nodulaire. Les deux souches de Rhizobium, utilisées pour l’inoculation des plantes, ont significativement augmenté la biomasse nodulaire, mais n’ont amélioré que légèrement la quantité d’azote fixée, et seulement en saison sèche. Indépendamment de la station et du traitement, la diminution de la pluviométrie d’une saison à l’autre est accompagnée, d’une part, d’une chute importante de la production des matières sèches aérienne et racinaire et, d’autre part, d’une amélioration très significative de la nodulation, tout particulièrement à Oued Béja. Cependant, les nodules développés dans les conditions de faible disponibilité de l’eau ont été mis en relation avec une très forte baisse des quantités d’azote fixées. En revanche, c’est l’apport d’azote minéral au sol qui a conduit à un enrichissement des parties aériennes en azote dans les stations, notamment en saison sèche.
Abstract
Effect of nitrogen fertilization, inoculation with Rhizobium sp. and precipitation on the biomass production and nitrogen content of chickpea. Water availability and soil fertility (especially soil N content) are the major factors affecting chickpea yield in Mediterranean regions, characterized by low rainfall, poorly distributed and rapid mineralization of organic matter. In order to study the effect of N-fertilization and inoculation with Rhizobium on chickpea biomass production and N-content, field experiment were carried out at two stations of INRAT in northern Tunisia during two consecutive years, 2000-2001 and 2001-2002. Results showed that N-fertilization significantly increased shoot biomass and N-content while decreasing nodule mass. Inoculation with both the two Rhizobium strains resulted in nodule mass increase but the shoot N-content was not significantly affected. Regardless of the stations and treatments, rainfall decrease between the two years of the experiment significantly reduced shoot and root biomass while improving nodulation, especially at Oued Beja. However, the nodule developed under water deficit conditions was correlated with a very high decrease of the nitrogen fixed. Nevertheless, the mineral nitrogen apply in soil improved the N-content of aerial parts in the two stations, especially in waterless season.
Table des matières
I. Introduction
1La nutrition azotée des légumineuses est d’autant plus difficile à étudier qu’elle résulte de la combinaison de deux voies très différentes, la fixation de l’azote atmosphérique et l’assimilation de l’azote minéral. La contribution de chacune de ces deux voies à l’alimentation en azote de la plante varie beaucoup selon les espèces et les conditions de la culture (Wery, 1987). L’amélioration de la production des légumineuses et leur utilisation en tant que précédent cultural nécessite donc de mieux connaitre les relations entre l’assimilation et la fixation de l’azote, d’une part, et l’influence des facteurs du milieu et des techniques culturales sur ces deux voies, d’autre part.
2Par ailleurs, la salinité et la sècheresse constituent des contraintes majeures limitant considérablement la productivité végétale sur une superficie d’environ 40 % de la surface terrestre, notamment en régions méditerranéennes (FAO, 1988). La tolérance des végétaux à ces contraintes est un phénomène complexe qui implique des particularités morphologiques et développementales avec des mécanismes physiologiques et biochimiques variés. Chez les légumineuses, leurs effets délétères s’exercent non seulement sur la plante hôte, mais aussi sur la croissance et la survie des populations du Rhizobium du sol (Singloton et al., 1982), sur le développement et le fonctionnement des nodosités et, donc, sur la capacité fixatrice d’azote de l’association symbiotique. La survie et la croissance des légumineuses dans ces conditions sont liées à des processus adaptatifs qui impliquent le transport et la compartimentation des ions, la biosynthèse et l’accumulation d’osmolytes organiques participant à l’ajustement osmotique et à des remaniements protéiques nécessaires au maintien de l’intégrité cellulaire (Sharma et al., 1992).
3En Tunisie, où les zones arides et semi-arides représentent 90 % de la surface totale, la culture du pois chiche endémique reste aléatoire en raison de la sècheresse et de la salinité des eaux et des sols. Le pois chiche est l’une des légumineuses les plus cultivées dans le monde pour l’alimentation humaine. En Tunisie, les superficies destinées à cette culture varient de 20 000 à 40 000 ha selon les années, ce qui correspond à environ 45 % de la surface totale des légumineuses à graines cultivées. C’est aussi une culture de qualité qui participe à l’amélioration des parcours culturaux. En effet, l’association des légumineuses avec les bactéries symbiotiques enrichit le sol en composés azotés pouvant être mis à profit par les cultures suivantes (Long, 1989). Bien que Rhizobium sp. soit utilisé en agriculture depuis près d’un siècle et que de nombreux travaux ont été consacrés à son étude dans les sols, les résultats acquis sont le plus souvent difficiles à interpréter et fréquemment contradictoires. À l’état libre ou associées aux légumineuses, ces bactéries sont, en effet, influencées par l’environnement (pH, température, oxygène dissous, humidité du sol) (Bekki et al., 1987 ; Fougère et al., 1991).
4Le métabolisme azoté et la synthèse protéique sont très perturbés par le stress osmotique qui affecte à la fois les populations rhizobiennes, la plante hôte et la relation symbiotique. L’initiation nodulaire est extrêmement sensible à la contrainte osmotique en raison de la réduction des sites d’infection de la racine, du nombre de poils racinaires et de la proportion de ceux qui portent les cordons d’infection (Zahran et al., 1986). Toujours en réponse à cette contrainte, un épaississement important des cortex nodulaires externe et interne est observé chez la luzerne. Parallèlement, les cellules non infectées contiennent de très nombreux amyloplastes, alors que les nodules développés en présence de NaCl en sont totalement dépourvus (Le Rudulier et al., 1995).
5Les analyses effectuées au cours des dernières années à propos de l’effet des contraintes de l’environnement sur la croissance végétale sous climat méditerranéen indiquent que l’azote pourrait être l’un des facteurs les plus limitants de la production végétale et bien qu’il soit possible d’augmenter la productivité de certains sols au moyen d’engrais azotés, les faibles bénéfices économiques qui en résultent font que la fertilisation est impossible dans beaucoup de conditions. C’est le cas, notamment, des environnements les plus arides où la réponse de la plante est limitée par la faible disponibilité de l’eau durant une longue période de son cycle de vie. En raison de ce type de limitation, une attention particulière doit être accordée à la fixation biologique de l’azote au moyen de l’utilisation d’associations légumineuses-Rhizobium (Johnson et al., 1981; Beck et al., 1988; Sanchez-Diaz et al., 1991).
6L’objectif du présent travail est d’étudier l’effet de l’inoculation par des bactéries symbiotiques, de la fertilisation azotée et du régime des pluies sur la production de biomasse et la teneur en azote du pois chiche dans deux régions différentes de la Tunisie.
2. Matériel et méthodes
2.1. Sites et conditions de l’expérimentation
7Les essais ont été conduits au cours des deux campagnes agricoles 2000-2001 et 2001-2002 caractérisées par une pluviométrie contrastée dans deux régions différentes du nord de la Tunisie. Il s’agit de deux stations de l’INRAT, l’une à Oued Béja (OB), gouvernorat de Béja (latitude 36,44 °N, longitude 9,11 °E) et l’autre à Oued Méliz (OM), gouvernorat de Jendouba (latitude 36,28 °N, longitude 8,29 °E). Dans ces deux stations, le climat est du type semi-aride tempéré, la température moyenne est de 19 °C et le sol est caractérisé par une texture argileuse. La saison pluvieuse s’étend du mois de septembre au mois de juin, avec des pluviométries annuelles de 550,9 mm en 2000-2001 et de 269 mm en 2001-2002, dans la station OB, et de 463,3 mm en 2000-2001 et 256,4 mm en 2001-2002, dans la station OM. Les teneurs en phosphore du sol sont de 32 et 39 ppm et celles en azote total de 2,77 et 2,18 %, respectivement dans les stations OB et OM.
2.2. Conduite de la culture
8La culture a été conduite chaque année et dans chaque station dans une parcelle expérimentale de 286 m2 de superficie, divisée en 16 parcelles élémentaires de 10 m2 séparées entre elles par une allée de 1 m. L’essai comporte quatre traitements répétés quatre fois en blocs complètement aléatoires. Le premier traitement constitué de plantes non inoculées est considéré comme témoin. Le second et le troisième traitement consistent en une inoculation avec Rhizobium sp., respectivement avec la souche de référence Rca 43 (ICARDA, Syrie) et la souche locale Rca 41 (INRAT, Tunisie), choisies dans la rhizothèque du Laboratoire des Grandes Cultures, Programmes de Légumineuses à Graines (INRAT), pour leurs performances symbiotiques. Le quatrième traitement consiste en un apport d’azote sous forme de nitrate d’ammonium, 33,5 % à raison de 60 kg d’azote par hectare dans leur milieu. La variété d’automne de pois chiche, INRAT 93.1, a été utilisée dans les essais. Un apport de superphosphates, 45 %, à raison de 90 kg du phosphate (P2O5) par hectare, est effectué au semis pour tous les traitements. Le désherbage et le binage sont effectués manuellement. L'inoculation des semences est effectué par enrobage au semis à raison de 4 g d’inoculum par kg de semences (FAO, 1988). L’inoculum utilisé contient plus de 108 bactéries par gramme. Pour éviter toute contamination, les parcelles élémentaires témoins et azotées sont préparées avant les parcelles inoculées.
2.3. Paramètres analysés
9Au stade floraison, 16 plantes par traitement ont été récoltées manuellement et séparées en parties aériennes, racines et nodules. Les racines sont rapidement rincées à l’eau puis essorées avec du papier filtre. Les nodules sont détachés de leurs racines, les masses de matière sèche des différents organes de la plante sont déterminées par pesée après dessiccation à l’étuve à 80 °C pendant 72 h. Cent mg de matière sèche finement broyée de la partie aérienne de la plante ont été utilisés pour le dosage d’azote par la méthode de Kjeldahl. Les analyses de variance et les erreurs standards des moyennes ont été réalisées pour déterminer la signification (p < 0,05) des différences entre les traitements.
3. Résultats et discussion
3.1. Croissance des parties aériennes et racinaires
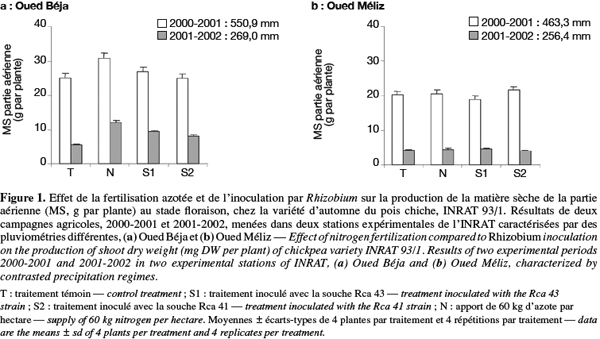
10Les résultats obtenus à la station Oued Béja (OB) ont montré que le traitement azoté a augmenté la biomasse aérienne par comparaison avec le témoin, sous les deux régimes de pluies (Figure 1a). Il apparait ainsi un effet favorable du nitrate (starter) sur la croissance des plantes qui s’est exprimé par une meilleure production de la matière sèche aérienne. Cet effet s’est exercé surtout à un stade précoce du développement de la plante (25 jours après la levée), où les nodules n’ont pas encore atteint la maturité et n’ont pas commencé la fixation de l’azote. Par ailleurs, les traitements d’inoculation qui n’ont pas abouti à des effets significatifs en 2001 (année pluvieuse) ont conduit en 2002 (année moins pluvieuse) à une augmentation des biomasses aériennes par rapport au témoin (Figure 1a). Par contre, les résultats obtenus dans la station Oued Méliz (OM) ont montré de légères variations de la biomasse aérienne en réponse aux différents traitements, mais qui ne sont pas significatives et ce, pour les deux régimes de pluies (Figure 1b).
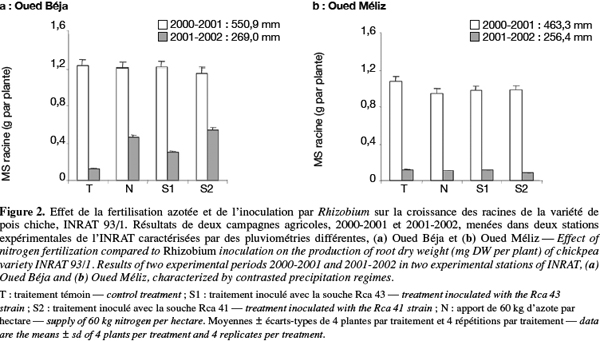
11Au niveau des racines, l’effet le plus remarquable sur la biomasse sèche reste celui du régime des pluies dans la station OB comme dans celle de OM (Figure 2). Quant aux effets des traitements, les seules différences significatives sont observées à OB et seulement durant l’année marquée par un faible régime des pluies (Figure 2a). En effet, une amélioration significative de la biomasse racinaire est observée en réponse à l’apport d’azote et à l’inoculation par les bactéries S1 et surtout S2, par comparaison avec le traitement témoin.
12On retiendra de cette analyse les résultats essentiels suivants :
13– Un effet positif de l’apport d’azote sur les biomasses aérienne et radiculaire de la plante est obtenu en année sèche dans la station OB, qui peut être attribué à l’enrichissement du sol en nitrate. Cela peut s’expliquer par le fait que, selon Salsac et al. (1984), l’azote pourrait jouer un rôle important dans l’ajustement osmotique vu le faible cout énergétique de son accumulation. De ce fait, un niveau d’accumulation élevé de cette forme d’azote devrait favoriser la capacité d’ajustement osmotique et la tolérance au déficit hydrique des légumineuses ;
14– L’inoculation par des bactéries fixatrices d’azote n’a pas amélioré la croissance des parties aériennes, ni celle des racines, par comparaison avec l’apport d’azote. Elle semble même l’avoir atténué ;
15– Dans la station OM, la croissance des parties aériennes comme celle des racines est indépendante des traitements. Il est possible que la richesse du sol de cette station soit à l’origine de l’absence de différences entre les traitements, sachant que l’apport d'azote minéral effectué est de 20,1 kg.ha-1 (60 kg.ha-1 de NH4NO3 33,5 %). Dans les essais au champ, un tel optimum est difficile à mettre en évidence car, dans la plupart des stations expérimentales, le sol est suffisamment pourvu en nitrate pour que le moindre apport de cet élément entraine un dépassement du niveau optimal (Wery, 1987) ;
16– Les rendements en biomasse enregistrés dans les deux stations sont nettement plus élevés en année humide qu’en année sèche. Autrement dit, la diminution de la pluviométrie d’une année à l’autre a entrainé une chute considérable de la production de matière sèche aérienne et racinaire (Figures 1 et 2). Ceci dénote le rôle prépondérant de l’alimentation hydrique dans l’élaboration de la matière sèche. Le déficit hydrique et l’irrégularité des pluies constituent, de ce fait, des facteurs limitants majeurs pour la production agricole en zone méditerranéenne.
3.2. Croissance nodulaire
17Les variations, en fonction des traitements, des masses de matière sèche nodulaire enregistrées dans les deux stations au terme des deux campagnes sont représentées sur les figures 3a et 3b. Il apparait, d’une part, que la culture sur sol témoin non inoculé a initié la formation de nodules sur les racines des plantes des deux stations et sous les deux régimes pluviométriques. D’autre part, la croissance de ces nodules semble dépendante des conditions d’humidité, mais seulement dans la station OB. En effet, la biomasse nodulaire sèche produite dans cette station sous régime pluviométrique limité est trois fois plus importante que celle produite sous régime humide. Toutefois, dans la station OM, les biomasses produites sous les deux régimes sont comparables. Par comparaison avec le traitement témoin, le traitement azoté a entrainé une inhibition de la formation et de la croissance nodulaire plus ou moins importante selon la station et le régime des pluies. Cet effet, peu marqué sous régime pluviométrique humide et dans les deux stations, est significativement plus important sous régime faible, mais seulement dans la station OB. Ceci pourrait s’expliquer par un effet direct du nitrate sur le processus d’infection et sur le fonctionnement des nodules (Drevon, 1984). Dans l’ensemble, ces résultats suggèrent l’existence de populations indigènes de bactéries symbiotiques dans les sols des deux stations et, qu’au moins dans la station OB, leur capacité d’infecter et de noduler les racines du pois chiche est fortement stimulée sous régime pluviométrique faible. En outre, cet effet est atténué par l’apport de nitrate au sol.
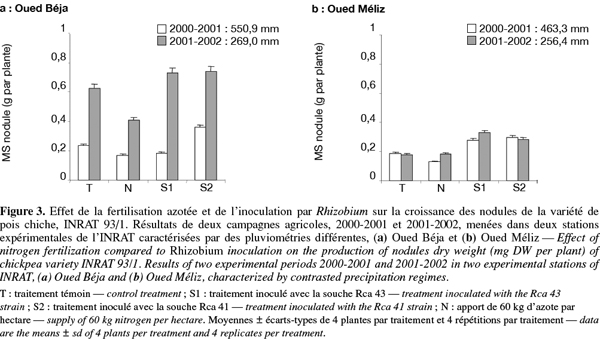
18En ce qui concerne les traitements d’inoculation, ils ont conduit à une amélioration modeste de la croissance nodulaire, par comparaison avec celle notée sur le sol témoin, et dont l’importance est dépendante de la station et du régime des pluies. Dans la station OB, des effets comparables des deux souches sont observés dans les conditions engendrées par un régime hydrique faible, alors que des différences significatives sont enregistrées entre les effets des deux bactéries d’inoculation dans les conditions relativement pluvieuses. Ces différences traduisent une meilleure efficacité des bactéries S2 dans ces conditions, par comparaison avec les bactéries S1 qui n’ont pas manifesté d’effet sur la croissance nodulaire.
19L’infectivité de la souche S1 est diminuée probablement en raison des conditions pédoclimatiques (type de sol, température et pluviométrie) qui sont tributaires de la région et de la saison (Evans, 1982 ; Heichel, 1987). Dans la station OM, les effets positifs des traitements d’inoculation sur la nodulation sont similaires et très peu dépendants du régime hydrique et de l’espèce bactérienne.
20En définitive, on retiendra que la production nodulaire à OB est supérieure à celle de OM, notamment dans le cas du régime de pluies le plus faible. Le type de sol peut être à l’origine de telles différences, comme il a été suggéré par Tellawi et al. (1986) qui ont constaté que ce facteur affecte significativement le nombre, la biomasse des nodules et l’absorption de l’azote. Il est aussi remarquable que la baisse de la pluviométrie en saison sèche n’a pas modifié la biomasse nodulaire dans la station OM, alors qu’elle l’a fortement augmentée dans la station OB. Cet effet positif de la sècheresse sur la croissance nodulaire serait le résultat d’une plus grande allocation de photosynthétats aux nodules, selon une étude comparative des effets de la sècheresse sur deux modèles de nodulation de deux cultivars de soja (Purcell et al., 1997). Dans un travail similaire sur le dolique, Figueiredo et al. (1998) ont pu démontrer une efficacité de la souche d’inoculation dans de telles conditions qu’ils ont attribuées au contenu en leghémoglobine des nodules, au potentiel hydrique foliaire et au poids de matière sèche des racines. Des travaux effectués auparavant sur le soja (Sprent, 1981) ont souligné l’importance de la taille des nodules dans leur capacité à supporter les conditions sèches. Selon ces travaux, les nodules de petite taille perdent rapidement leur réserve en eau, alors que ceux de taille plus grande possèdent un parenchyme cortical plus épais qui empêche ou réduit leur déshydratation lorsque la culture est réalisée sur sol sec. Il apparait ainsi que les nodules volumineux s’adaptent bien aux conditions du stress hydrique, puisque les cellules microbiennes, y compris le Rhizobium, sont capables de résister à des potentiels hydriques plus bas que la plupart des cellules des plantes supérieures. Par conséquent, il est probable que depuis l’infection par le Rhizobium jusqu’au fonctionnement des nodules différenciés, les facteurs les plus importants qui limitent la fixation dans des conditions de stress hydrique résident dans la plante hôte (Sanchez-Diaz et al., 1995).
3.3. Taux et quantité d’azote total de la partie aérienne
21Les variations du taux et de la quantité d’azote total de la partie aérienne dans les deux stations sont représentées en fonction des traitements, respectivement à la figure 4 et à la figure 5. La culture sur sol témoin non inoculé a conduit à des niveaux d’accumulation en azote voisins et légèrement supérieurs à 3 %, et ce indépendamment du régime pluviométrique et de la station de culture (Figure 4). L’effet de l’apport d’azote sur ces niveaux s’est traduit par des variations significatives dans la station OB, indiquant une baisse en année humide et une augmentation en année sèche. Toutefois, les niveaux enregistrés chez les plantes de la station OM sont très peu ou pas différents de ceux du traitement témoin.
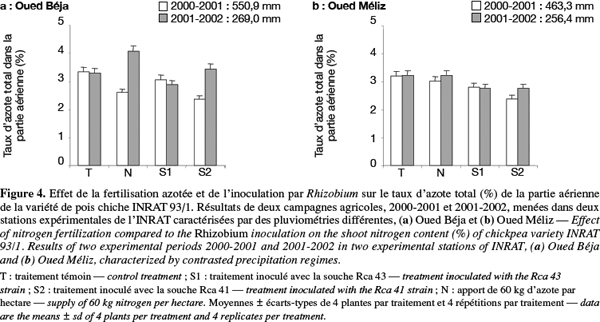
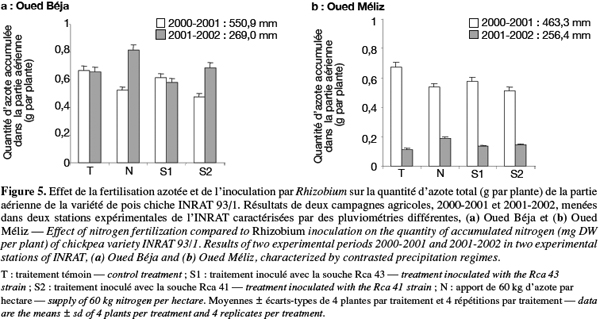
22Par ailleurs, les variations induites par l’inoculation avec deux souches de bactéries symbiotiques, S1 et S2, semblent dépendre surtout de la souche et du régime de pluies (Figure 4). Par comparaison avec celle du traitement témoin, l’accumulation d’azote dans les parties aériennes est légèrement abaissée en réponse à l’inoculation avec la souche S1, et ce quels que soient le régime pluviométrique et la station. En revanche, une baisse plus marquée du niveau d’accumulation de l'azote est notée lorsque l’inoculation est faite avec l’autre souche bactérienne (S2), notamment sous régime pluvieux des deux stations. Dans les conditions moins pluvieuses, cet effet n’est observé que dans la station OM et il est relativement atténué.
23En ce qui concerne les quantités d’azote, les résultats de la figure 5 ne tiennent compte que de la fraction allouée aux parties aériennes. Celles distribuées aux racines et aux nodules n’étant pas déterminées, nous avons supposé qu’elles sont très faibles étant donné les faibles biomasses des ces organes par comparaison avec celles des organes aériens. L’examen de cette figure révèle que dans chacune des deux stations, les quantités d’azote alloué aux parties aériennes sont fortement tributaires de l’apport d’eau : indépendamment de la station et des traitements, les parties aériennes demeurent plus riches en azote en saison humide qu’en saison sèche, et ce en dépit d’une plus faible biomasse nodulaire (efficacité des nodules). Ceci suggère que les nodules développés en saison humide expriment une meilleure capacité fixatrice d’azote, tout en étant moins nombreux.
24Par ailleurs, l’apport d’ammonitrate au sol a conduit à une allocation supplémentaire de l’azote aux parties aériennes dans les conditions humides de la station OB, et par contre, à une moindre allocation dans celles de la station OM. En revanche, le même traitement a toujours conduit, sous régime pluvieux sec, à un enrichissement en azote de ces organes : les quantités accumulées représentent environ le double de celles du traitement témoin dans les deux stations.
25Nos résultats ont montré aussi que, par comparaison avec le traitement témoin, l’inoculation avec les deux souches rhizobiales, notamment avec la souche S2, a entrainé une baisse significative des quantités d’azote dans les parties aériennes en saison humide et à une augmentation très peu marquée, mais significative, en saison sèche dans chacune des deux stations (Figure 5). Toutefois, cet effet bénéfique de l’inoculation sur la fixation de l’azote demeure très modeste par comparaison à celui enregistré en réponse à un apport d’ammonitrate. Ces résultats suggèrent que les bactéries symbiotiques utilisées dans cette étude n’ont pas exprimé leurs potentialités réelles de fixation de l’azote moléculaire, éventuellement en raison d’une forte compétition avec les populations bactériennes indigènes du sol.
26Enfin, les variations des quantités d’azote sont généralement similaires à celles de la croissance (Figure 1), ce qui permet de suggérer qu’elles soient l’expression d’un effet indirect de l’apport d’eau sur la croissance des plantes.
4. Conclusion
27La fertilisation azotée a entrainé une augmentation de la biomasse aérienne et une diminution de la biomasse nodulaire. Ceci confirme l’effet du nitrate sur le processus d’infection et sur le fonctionnement des nodosités. De plus, les deux souches de Rhizobium, S1 et S2, utilisées pour l’inoculation des plantes ont significativement augmenté la biomasse nodulaire, mais n’ont amélioré que légèrement la quantité d’azote fixée et seulement en saison sèche.
28Indépendamment de la station et du traitement, la variation de la pluviométrie d’une saison à l’autre est accompagnée, d’une part, d’une chute importante de la production de matières sèches aérienne et racinaire et, d’autre part, d’une amélioration très significative de la nodulation, tout particulièrement à OB. Il semble bien que les processus d’initiation et de formation nodulaires se sont mieux exprimés en conditions de déficit hydrique. Cependant, les nodules développés dans ces conditions de faible disponibilité de l’eau n’ont pas permis une meilleure fixation de l’azote, mais c’est plutôt une très forte baisse des quantités fixées qui a été enregistrée. En revanche, c’est l’apport d’azote minéral au sol, sous forme d’ammonitrate, qui a conduit à un enrichissement des parties aériennes en azote dans les stations, notamment en saison sèche.
29Etant donné les aléas de la pluviosité en zone méditerranéenne en général et en Tunisie en particulier, il serait souhaitable d’approfondir les recherches sur la nodulation et sur la fixation symbiotique de l’azote en condition de déficit hydrique afin de sélectionner des symbioses fixatrices pour le maintien d’une bonne production agricole et d’une quantité d’azote appréciable au niveau du sol. L’amélioration de la fixation azotée pourrait être basée sur des populations natives. Des études devraient être conduites pour améliorer la capacité de nodulation de ces bactéries et une meilleure expression de leur potentiel de fixation de l’azote. La différence des caractères symbiotiques entre les populations natives des différents sites mérite d’être analysée en rapport avec la variabilité des conditions climatiques et édaphiques.
Bibliographie
Beck D.P. & Materon L.A., 1988. Nitrogen fixation by legumes in Mediterranean agriculture. Dordrecht, The Netherlands: Martinus Nijhoff Publishers.
Bekki A., Trinchant J.C. & Rigaud J., 1987. Nitrogen fixation (C2H2 reduction) by Medicago nodules and bacteroids under sodium chloride stress. Physiol. Plant., 71, 61-67.
Drevon J.J., 1984. Etude du métabolisme de l’hydrogène dans la symbiose Phaseoleae-rhizobium. Thèse de doctorat : Université des Sciences et Techniques du Languedoc (USTL), Montpellier (France).
Evans J., 1982. Symbiosis, nitrogen and dry matter distribution in chickpea (Cicier arietinum). Exp. Agric., 18, 339-351.
FAO, 1988. Programme de coopération technique. Programme de développement des productions fourragères et de l'élevage. Rapport de synthèse. Rome : FAO.
Figueiredo M.V.B., Burity H.A. & De França F.P., 1998. Water deficit stress effects on N2 fixation in cowpea inoculated with different Bradyrhizobium strains. Can. J. Plant Sci., 38, 311-321.
Fougère F., Le Rudulier D. & Streeter J.G., 1991. Effects of salt stress on amino acid, organic acid and carbohydrate composition of roots, bacteroids and cytosol of alfalfa (Medicago sativa L.). Plant Physiol., 96, 1228-1236.
Heichel G., 1987. Legume nitrogen: symbiotic fixation and recovery by subsequent crops. In: Helsel Z.R., ed. Energy in plant nutrition and best control. Amsterdam: Elsevier, 63-80.
Johnson D.A., Rumbaugh M.D. & Asay K.H., 1981. Plant improvement for semi-aride rangelands: possibilities for drought resistance and nitrogen fixation. In: Montieth J. & Web C., eds. Soil water and nitrogen in Mediterranean-type environments. The Hague, The Netherlands: Martinus Nijhoff/Dr. W. Junk Publishers, 279-303.
Le Rudulier D., Gloux K., Poggi M.C. & Noël J.P., 1995. Accumulation, biosynthesis and fate of proline betaine in nodulated alfalfa plants (Medicago sativa L.) subjected to salinity stress. In: Leigh R.A. & Blake-Kalff M.M.A., eds. Proceedings of the 2nd Stress net conference, Salsomaggiore, European Commission VI, 257-62.
Long S.R., 1989. Rhizobium-legume nodulation: life together in the underground. Cell, 56, 203-214.
Purcell L.C., De Silva M., King C.A. & Kim W.H., 1997. Biomass accumulation and allocation in soybean associated with genotypic differences in tolerance of nitrogen fixation to water deficits. Plant Soil, 196, 101-113.
Salsac L. & Chaillou S., 1984. Nutrition azotée des végétaux : importance physiologique et écologique de la fourniture d’azote sous forme nitrique ou ammoniacal. Bull. Soc. Ecophysiol., 9, 111-128.
Sanchez-Diaz M. & Aguirreolea J., 1991. Fijacion de nitrogeno en embeintes àriodos y semiàriodos. In: Megias Guijo M., Palomares Diaz A.J. & Ruis Berraquero F., eds. Aportaciones a la biologia de la fijacion del nitrogeno atmosférico. Sevilla, Spain: Publicaciones Universidad de Sevilla.
Sanchez-Diaz M., Aguirreolea J., Goicochea N. & Antolin M.C., 1995. Limitation de la fixation symbiotique d’azote et autres aspects physiologiques des légumineuses des zones méditerranéennes. In : Actes du colloque, Facteurs limitant la fixation symbiotique de l’azote dans le bassin méditerranéen, 6-8 avril 1994, Montpellier, France. Paris : INRA.
Sharma S.K. & Kumar S., 1992. Effect of salinity on Na+ and Cl- content in different nodule organs of chickpea and the basis on ion expression. Biol. Plant., 34, 311-317.
Singleton P.W., Swaify A.E. & Bohlool B.B., 1982. Effect of salinity on Rhizobium growth and survival. Appl. Environ. Microbiol., 44, 884-890.
Sprent J.I., 1981. Nitrogen fixation. In: Paleg L.G. & Aspinall D., eds. The physiology and biochimistry of drought resistance in plants. Sydney, Australia: Academic Press, 131-151.
Tellawi A., Haddad N. & Hattar B., 1986. Effect of several Rhizobium strains on nodulation, nitrogen uptake and yield of chick peas (Cicer arietinum L.). Z. Pflanzenernaehr. Bodenkd., 149, 314-322.
Wery J., 1987. Relation entre la nutrition azotée et la production chez les légumineuses. In : Actes du colloque, Nutrition azotée des légumineuses, 19-21 novembre 1985, Versailles, France. Paris : INRA.
Zahran H.H. & Sprent J.L., 1986. Effects of sodium chloride and polyethylene glycol on root-hair infection and nodulation of Vicia faba L. plants by Rhizobium leguminosarum. Planta, 167, 303-309.
Pour citer cet article
A propos de : Boulbaba L’taief
Université de Tunis El Manar. Faculté des Sciences. Département de Biologie. Campus universitaire. TU-1060 Tunis (Tunisie) – Institut National de la Recherche Agronomique de Tunisie (INRAT). Laboratoire des Grandes Cultures. Rue Hédi Karray. TU-2080 Ariana (Tunisie). E-mail : lboulaba@yahoo.com
A propos de : Bouaziz Sifi
Institut National de la Recherche Agronomique de Tunisie (INRAT). Laboratoire des Grandes Cultures. Rue Hédi Karray. TU-2080 Ariana (Tunisie).
A propos de : Mainassara Zaman-Allah
Institut National de la Recherche Agronomique de Tunisie (INRAT). Laboratoire des Grandes Cultures. Rue Hédi Karray. TU-2080 Ariana (Tunisie).
A propos de : Mokhtar Hajji
Université de Tunis El Manar. Faculté des Sciences. Département de Biologie. Campus universitaire. TU-1060 Tunis (Tunisie).
A propos de : Mokhtar Lachaâl
Université de Tunis El Manar. Faculté des Sciences. Département de Biologie. Campus universitaire. TU-1060 Tunis (Tunisie).
