


- Accueil
- volume 13 (2009)
- numéro 3
- Effet de la bactérisation des graines sur la croissance des plants de Cedrus atlantica Manetti


Visualisation(s): 4306 (31 ULiège)
Téléchargement(s): 289 (3 ULiège)

Effet de la bactérisation des graines sur la croissance des plants de Cedrus atlantica Manetti

Notes de la rédaction
Reçu le 15 mai 2008, accepté le 21 octobre 2008
Résumé
Cinq souches rhizobactériennes ont été testées pour leur effet sur la croissance des plants de Cedrus atlantica Manetti en pépinière forestière avant transplantation sur le terrain. Les résultats obtenus ont montré un effet significatif des souches bactériennes sur la croissance de semis de cèdre et seules les deux Pseudomonas fluorescens A6RI et TGI252 ont stimulé significativement la longueur de la tige, le diamètre au collet, le poids sec des racines et le nombre de racines courtes. Cela nous permettra d'envisager la mise au point de cette technique pour la production de plants de qualité.
Abstract
Bacterization effect of seeds on the growth of Cedrus atlantica Manetti plants. The beneficial effect of five rhizobacterial strains on the growth of Cedrus atlantica plants was evaluated at forest nursery before out-planting. The obtained results showed a significant effect of bacterial strains on Cedar seedlings growth and only Pseudomonas fluorescens A6RI and TGI252 significantly increased stem length, neck diameter, root dry weight and number of root tips. It will allow us to adapt this technology for the production of quality plants.
Table des matières
1. Introduction
1Au Maroc, Cedrus atlantica Manetti constitue les principales formations forestières montagnardes, sur environ 132 000 ha. Ces peuplements présentent d'importants atouts en rapport avec leur grande diversité biologique et leur impact sur l'équilibre socio-économique des zones de montagne et de plaine. Cependant, la cédraie marocaine fait face à de nombreuses pressions telles que la surexploitation et les agressions parasitaires. En plus, les programmes de reconstitution de la cédraie sont confrontés à un taux de réussite inférieur à 50 %. Ceci est principalement dû à la qualité des plants de cèdre produits en pépinière forestière qui souffrent généralement d'une faible croissance et d'un système racinaire réduit (Mhirit, 1994 ; Mhirit et al., 1999).
2Les inoculums bactériens ont été introduits dans le sol afin d'améliorer la croissance et la santé des plants tout en réduisant l'utilisation des produits chimiques (Schipper et al., 1995 ; Gamalero et al., 2002). Les groupes de bactéries utilisées appartiennent notamment aux genres Bacillus (Holl et al., 1988), Pseudomonas (O'Neill et al., 1992) et Enterobacter (Beall et al., 1989). Cet effet bénéfique des bactéries sur la croissance des plantes est souvent évalué par l'augmentation du poids des racines et des tiges.
3Cependant, les travaux menés sur les bactéries stimulant la croissance des semis d'arbres sont quasi-absentes et à notre connaissance, aucune étude n'a été effectuée sur l'amélioration de la croissance des semis de cèdre. En plus, il s'est avéré que l'utilisation des souches bactériennes promotrices de la croissance des semis d'arbres en pépinière forestière est très intéressante et ceci est dû d'une part aux résultats positifs obtenus en agriculture et en horticulture et d'autre part, au faible cout et à la facilité d'application des opérations d'inoculations bactériennes en pépinière (Chanway, 1997 ; Vonderwell et al., 2000).
4Dans ce travail, nous avons évalué l'influence de cinq souches bactériennes appartenant aux espèces Pseudomonas fluorescens, Lactococcus lactis et Paenibacillus polymyxa sur les paramètres de croissance des plants de Cedrus atlantica en pépinière forestière, en l'occurrence le diamètre au collet, la longueur de la tige, le poids sec racinaire et le nombre de racines courtes.
2. Matériel et méthodes
2.1. Souches bactériennes
5Les souches bactériennes utilisées dans ce travail nous ont été fournies par l'Unité Mixte de Recherche (UMR), Interactions Arbre/Micro-organismes (IAM), INRA, Champenoux, France.
6La souche de Pseudomonas fluorescens 92 a été isolée à partir d'un carpophore du champignon ectomycorhizien Suillus grevillei (Varese et al., 1996). La souche Pseudomonas fluorescens A6RI est un mutant résistant à la rifampicine et elle est conservée dans la collection française de bactéries phytopathogènes (CFBP, Angers, France) sous le numéro CFBP2392 (Gamalero et al., 2002 ; 2004). La souche TGI252 a été isolée à partir d'un carpophore de Tricholoma cedrorum provenant de la rhizosphère d'une cédraie du Moyen Atlas au Maroc et elle a été identifiée comme Pseudomonas fluorescens biotype G par la méthode BIOLOG. La souche Paenibacillus polymyxa a été isolée pour la première fois à partir de la rhizosphère du blé et de l'orge (Lindberg et al., 1984), ensuite elle a été caractérisée dans la rhizosphère du pin (Chanway et al., 1991 ; Shishido et al., 1996), elle est connue sous le code B7. La souche Lactococcus lactis ssp. lactis est une bactérie communément isolée à partir des produits végétaux et laitiers, elle est connue sous le code MR3.
2.2. Préparation des graines
7Les graines de cèdre originaires de la région de Sehb (Moyen Atlas, Maroc) ont été rapportées de la station régionale des semences d'Azrou (Moyen Atlas, Maroc). Les graines ont été stérilisées en surface par trempage dans de l'eau oxygénée à 33 % pendant 5 min. Après trois lavages à l'eau distillée stérile, les graines ont été stockées dans le réfrigérateur à + 4 °C pendant deux semaines.
2.3. Conditions de culture
8L'expérimentation a été menée dans des portoirs rigides de 38 alvéoles de 400 ml chacune, préalablement désinfectés par trempage dans un bain d'hypochlorite de sodium (5 %). Le substrat de remplissage, non stérilisé, était celui habituellement utilisé dans la pépinière forestière d'Azrou (Moyen Atlas) pour la culture des plants. Celui-ci est constitué d'un mélange en quantités égales de terreau forestier, de sable et de compost. Les portoirs ont été installés en surélévation sur des piquets en bois et en plein air pour assurer l'autocernage des racines. En général, les essais sont menés dans des conditions similaires à celles de la production des plants de cèdre dans la pépinière.
2.4. Préparation de l'inoculum bactérien et bactérisation des graines
9Les souches bactériennes ont été mises en culture séparément à 28 °C pendant 48 heures sur un milieu nutritif gélosé constitué de bouillon de soja tryptique 1/10. Ensuite, les bactéries ont été mises en suspension dans 10 ml du tampon MgSO4, 7H2O (0,1M). Chaque suspension bactérienne est lavée trois fois par centrifugation et le culot est resuspendu dans une quantité minimum du tampon MgSO4, 7H2O (0,1M). La densité de l'inoculum est ajustée à 1010 ufc.ml-1 en utilisant une courbe d'étalonnage établie à une densité optique de 600 nm.
10Les graines de cèdre stériles ont été trempées dans 50 ml de la suspension bactérienne (1010 ufc.ml-1) pendant une durée de 20 min. Des tests préliminaires ont montré que cette procédure d'inoculation permet d'atteindre une concentration bactérienne de 107 ufc par graine. Les graines témoins ont été immergées dans une solution de sulfate de magnésium pendant la même durée.
2.5. Conditions de cultures et mesures des paramètres de croissance
11Les graines traitées ou non par les souches bactériennes ont été semées à raison de 3 graines par alvéole. Dans le cas de germination multiple, un seul semis est laissé par alvéole, les autres sont éliminés. Trois portoirs de 38 pots chacun ont été utilisés par traitement (trois répétitions). L'arrosage s'effectue automatiquement deux fois par jour (le matin et le soir). A deux mois (T1), quatre mois (T2) et six mois (T3) après la date d'inoculation, cinq plants par répétition (15 plants par traitement) ont été prélevés, le diamètre au collet et la longueur de la tige ont été mesurés. Ensuite, les racines ont été lavées et leur poids sec ainsi que le nombre de toutes les racines courtes ont été quantifiés par comptage sous la loupe binoculaire.
2.6. Analyses statistiques
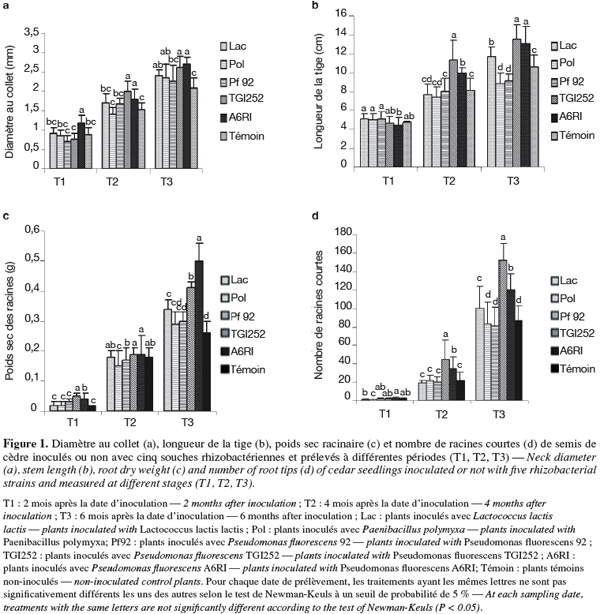
12Les valeurs ont été soumises à une analyse de la variance à l'aide du logiciel STAT-ITCF, Paris, France, 1987. La comparaison des moyennes a été effectuée par le test de Newman-Keuls (test de Fisher, figure 1) à un seuil de probabilité de 5 %.
3. Résultats et discussion
13Les semis de Cedrus atlantica inoculés avec la souche bactérienne P. fluorescens A6RI ont montré une augmentation hautement significative (p < 0,0000) par rapport au témoin, de leur diamètre au collet dès le deuxième mois d'inoculation T1 (Figure 1, a). A quatre mois d'inoculation (T2), P. fluorescens souche TGI252 a entrainé une amélioration hautement significative (p < 0,000) du diamètre au collet. Au temps d'échantillonnage T3, toutes les souches bactériennes ont provoqué une croissance positive du diamètre au collet des semis de Cedrus atlantica. Néanmoins, ce sont les deux souches bactériennes A6RI et TGI252 qui ont permis plus d'augmentation du diamètre au collet par rapport aux témoins, avec 29 et 26 % respectivement (Figure 1, a).
14Aucune souche n'a montré une influence significative sur la longueur de la tige des semis de cèdre âgés de deux mois (Figure 1, b). Après quatre mois d'inoculation (T2), les deux souches bactériennes TGI252 et A6RI ont stimulé significativement (p < 0,0000) la croissance en longueur des plants pour atteindre à six mois, des taux respectifs d'environ 27 % et 24 % supérieurs au témoin.
15A six mois, le poids sec des racines des traitements inoculés avec A6RI et TGI252 est significativement (p < 0,0000) supérieur de 92 % et 58 % respectivement par rapport au témoin non inoculé (Figure 1, c). Ceci montre que ces deux souches bactériennes stimulent aussi bien les parties aériennes que racinaires des plants de cèdre. Ce qui corrobore les résultats de plusieurs auteurs (Schönwitz et al., 1989 ; Lippmann et al., 1995) qui ont démontré que l'élévation de la biomasse de la partie racinaire est étroitement corrélée à l'augmentation de la biomasse de la partie aérienne chez les plants.
16Quant au nombre de racines courtes, aucune différence significative n'a été observée par rapport au témoin chez les plants inoculés par les cinq souches bactériennes jusqu'à deux mois (Figure 1, d). Néanmoins à partir de cette date, les deux souches de P. fluorescens A6RI et TGI252 ont significativement (p < 0,0000) augmenté le nombre de racines courtes par rapport au témoin non inoculé d'un facteur d'environ 106 % et 58 %, respectivement à quatre mois, et d'un facteur d'environ 87 % et 51 % respectivement à six mois.
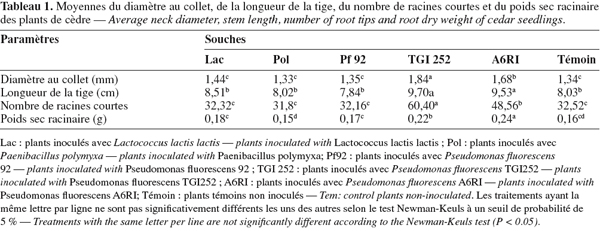
17Indépendamment de l'âge des plants, les résultats du tableau 1 confirment une influence positivement significative des deux souches bactériennes TGI252 et A6RI sur tous les paramètres de croissance en comparaison avec les autres bactéries et les témoins non-inoculés.
18Les deux souches A6RI et TGI252 ont stimulé la croissance du système racinaire (biomasse racinaire et nombre de racines courtes), permettant ainsi une plus grande exploration du sol et par conséquent, une augmentation de la surface d'échange et donc une nutrition améliorée, ce qui s'est traduit aussi par un meilleur développement de la partie aérienne. Il a été démontré que les systèmes racinaires les plus longs sont les plus adaptés à l'exploration du sol (Berta et al., 2002). La stimulation de la croissance du système racinaire des plants inoculés par A6RI et TGI252 pourrait être expliquée par le fait que ces deux souches bactériennes peuvent agir par l'intermédiaire d'hormones. En effet, Glick (1995) a démontré que la synthèse des auxines par les rhizobactéries est impliquée dans l'élongation racinaire. Il a été prouvé que la souche A6RI est capable de produire de l'acide indol-3-acétique (AIA) (Gamalero et al., 2002), ce qui pourrait expliquer son effet positif sur la croissance des plants de cèdre. Gamalero et al. (2002) ont souligné aussi que P. fluorescens A6RI agit positivement sur le développement du système racinaire de la tomate. Ceci démontre que cette souche est capable de promouvoir de manière non spécifique la croissance de différents plants. Ces mêmes auteurs (Gamalero et al., 2002) ont rapporté que la souche A6RI a augmenté de manière significative tous les paramètres morphologiques (volume, surface et biomasse) du système racinaire de la tomate, à l'exception du nombre de racines courtes. Au contraire, nos résultats ont montré une augmentation significative aussi bien de la biomasse racinaire que du nombre de racines courtes des plants de cèdre traités par cette souche. Ceci revêt une grande importance car les racines courtes sont le siège de la symbiose ectomycorhizienne. Par ailleurs, A6RI a été décrite comme agent de biocontrôle contre les maladies de la pourriture racinaire (Gamalero et al., 2002), ce qui pourrait influencer l'état sanitaire des racines des plants, surtout qu'ils sont cultivés sur substrat non stérile et donc colonisés par des champignons phytopathogènes. Plusieurs études ont signalé que certaines bactéries stimulatrices de la croissance des plants sont capables de produire des substances phytohormonales et des sidérophores ayant des actions antagonistes vis-à-vis des agents phytopathogènes (Nair et al., 1991 ; Ehteshmul-Haque et al., 1993 ; Yao et al., 2002).
19L'origine rhizosphérique de la souche TGI252 pourrait être aussi l'un des facteurs contribuant dans l'activité positive observée chez les plants de cèdre. Les mêmes résultats ont été rapportés par Gamalero et al. (2004) pour la souche P. fluorescens 190r, isolée à partir de la rhizosphère de la tomate. En effet, l'inoculation par la souche bactérienne 190r a provoqué l'accroissement des poids frais des tiges et des racines de la tomate. Chanway et al. (1993) ont signalé que la stimulation de la croissance des semis est statistiquement significative seulement quand l'écotype de Pseudotsuga menziesii a été inoculé avec la bactérie rhizosphérique avec laquelle il avait précédemment été en contact dans la nature.
20En ce qui concerne le diamètre au collet et le nombre de racines courtes, nos résultats ont montré également des différences significatives (P < 0,05) qui apparaissent tôt (dès le deuxième mois) entre les traitements inoculés par A6RI et les témoins des plants de cèdre. L'action de TGI252 sur ces paramètres apparait plus tard, à quatre mois et plus. Ceci est probablement dû à une différence entre le mécanisme d'action d'A6RI et de TGI252 ; l'action rapide de la souche A6RI pourrait être expliquée par un effet hormonal de type primaire qui agit directement sur la croissance des racines des plants ; par contre, l'action retardée de TGI252 suggère un effet nutritionnel par la bactérie qui mobilise en premier lieu les nutriments de la rhizosphère et ensuite le système racinaire de la plante, qui en profite d'une manière indirecte.
21Les deux souches bactériennes Paenibacillus polymyxa et P. fluorescens 92 ont inhibé de manière significative (p < 0,0000) la longueur de la tige des plants de cèdre âgés de six mois (Figure 1, a). Cette longueur, qui est en moyenne de 10,60 cm chez le témoin, a été réduite à 8,90 et 9,10 cm respectivement chez P. polymyxa et P. fluorescens 92. Cet effet antagoniste vis-à-vis de la croissance des plants a été déjà expliqué par des phénomènes de compétition nutritive quand la densité bactérienne est élevée (Kapulnik et al., 1995 ; Frey-Klett et al., 1999 ; Bending et al., 2002). Pourtant, ces deux bactéries ont été décrites respectivement par Gamalero et al. (2003) et Chanway et al. (1991) comme étant productrices d'AIA et capables de promouvoir la croissance des semis auxquels elles sont inoculées. Ceci laisse supposer que dans nos conditions expérimentales, il y avait soit un problème de spécificité ou une mauvaise colonisation racinaire par ces deux souches.
4. Conclusion
22Nos résultats apportent des données nouvelles sur l'effet bénéfique que peut avoir la bactérisation des graines avec des souches bactériennes sur la croissance de semis de cèdre en pépinière forestière. Les deux souches bactériennes utilisées A6RI et la TGI252 ont pu promouvoir la croissance des plants de cèdre ainsi que l'augmentation de leur nombre de racines courtes, site de formation des ectomycorhizes.
23Ce nouveau type de biotechnologie pourrait être développé et utilisé pour la culture et la production de plants de cèdre de qualité et par conséquent favoriser les facteurs qui contribuent à la réussite des opérations de reboisement au Maroc.
Bibliographie
Beall F. & Tipping B., 1989. Plant growth-promoting rhizobacteria in forestry. In: Proceedings of the forest research marketing. Abstract 117, Ontario Forest Research Committee, Toronto.
Bending G.D., Poole E.J., Whipps J.M. & Read D.J., 2002. Characterisation of bacteria from Pinus sylvestris-Suillus luteus mycorrhizas and their effects on root-fungus interactions and plant growth. FEMS Microbiol. Ecol., 39, 219-227.
Berta G., Fusconi A. & Hooker J.E., 2002. Arbuscular mycorrhizal modifications to plant root systems. In: Gianinazzi S. & Schuepp H., eds. Mycorrhizal technology: from genes to bioproducts achievement and hurdles in arbuscular mycorrhizal research. Basel, Switzerland: Birkhäuser, 71-101.
Chanway C.P., 1997. Inoculation of tree roots with plant growth promoting soil bacteria: an emerging technology for reforestation. Forest Sci., 43, 99-112.
Chanway C.P., Radley R.A. & Holl F.B., 1991. Inoculation of conifer seed with plant growth promoting Bacillus strains causes increased seedling emergence and biomass. Soil Biol. Biochem., 23, 575-580.
Chanway C.P. & Holl F.B., 1993. Ecological growth response specificity of two Douglas-fir ecotypes inoculated with coexistent beneficial rhizosphere bacteria. Can. J. Bot., 72, 582-586.
Ehteshamul-Haque S. & Ghaffar A., 1993. Use of rhizobia in the control of root rot diseases of sunflower, oak and munybean. J. Phytopathol., 138, 157-163.
Frey-Klett P., Churin J.L., Pierrat J.C. & Garbaye J., 1999. Dose effect in the dual inoculation of an ectomycorhizal fungus and a mycorrhiza helper bacterium in two forest nurseries. Soil Biol. Biochem., 31, 1555-1562.
Gamalero E. et al., 2002. Morphogenetic modifications induced by Pseudomonas fluorescens A6RI and Glomus mosseae BEG12 in the root system of tomato differ according to plant growth conditions. New Phytol., 155, 293-300.
Gamalero E. et al., 2003. Characterization of functional traits of two fluorescent pseudomonads isolated from basidiomes of ectomycorrhizal fungi. Soil Biol. Biochem., 35, 55-65.
Gamalero E. et al., 2004. Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal fungus on tomato plant growth, root architecture and P acquisition. Mycorrhiza, 14, 185-192.
Glick B.R., 1995. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol., 41, 109-117.
Holl F.B., Chanway C.P., Turkingon R. & Radley R., 1988. Growth response of crested wheatgrass (Agropyron cristatum L.), white clover (Trifolium repens L.) to inoculation with Bacillus polymixa. Soil Biol. Biochem., 20, 19-24.
Kapulnik Y., Okon Y. & Henis Y., 1995. Changes in root morphology of wheat caused by Azospirillum inoculation. Can. J. Microbiol., 31, 881-887.
Lindberg T. & Granhall U., 1984. Isolation and characterization of dinitrogen-fixing bacteria from the rhizosphere of temperate cereals and forage grasses. Appl. Environ. Microbiol., 48, 683-689.
Lippmann B., Leinhos V. & Bergmann H., 1995. Influence of auxin producing rhizobacteria on root morphology and nutrient accumulation of crops. I. Changes in root morphology and nutrient accumulation in maize (Zea mays L.) caused by inoculation with indole-3-acetic acid (IAA) producing Pseudomonas and Acinetobacter strains or IAA applied exogenously. Angew. Bot., 69, 31-36.
Mhirit O., 1994. Le cèdre de l'Atlas (Cedrus atlantica Manetti) : présentation générale et état des connaissances à travers le réseau Silva Mediterranea « le cèdre ». Ann. Rech. For. Maroc, 27, 4-21.
Mhirit O. et al., 1999. Le grand livre de la forêt marocaine. Sprimont, Belgique : Editions Mardaga.
Nair M.G., Safir G.R. & Siquiera J.O., 1991. Isolation and identification of vesicular-arbuscular mycorrhiza stimulatory compounds from Clover (Trifolium repens) roots. Appl. Environ. Microbiol., 57, 434-439.
O'Neill G.A., Radley R.A. & Chanway C.P., 1992. Variable effects of emergence-promoting rhizobacteria on conifer seedling growth under nursery conditions. Biol. Fertil. Soils, 13, 45-49.
Schipper B., Scheffer R.J., Lugtenberg B.J.J. & Weisbeck P.J., 1995. Biocoating of seeds whith plant growth-promoting rhizobacteria to improve plant establishment. Outlook Agric., 24, 179-185.
Schönwitz R. & Ziegler H., 1989. Interaction of maize roots and rhizosphere microorganisms. Z. Pflanzenern. Bodenk., 152, 217-222.
Shishido M., Paterson D.L., Massicote H.B. & Chanway C.P., 1996. Pine and spruce seedling growth and mycorrhizal infection after inoculation with plant growth promoting Pseudomonas strains. FEMS Microbiol. Ecol., 21, 109-119.
Varese G.C. et al., 1996. Bacteria associated with Suillus grevillei sporocarps and ectomycorrhizae and their effect on in vitro growth of the mycobiont. Symbiosis, 21, 129-147.
Vonderwell J.D. & Enebak S.A., 2000. Differential effects of rhizobacterial strain and dose on the ectomycorrhizal colonization of loblolly pine seedlings. For. Sci., 46(3), 411-437.
Yao M.K., Tweddell R.J. & Desilets H., 2002. Effect of two vesicular-arbuscular mycorrhizal fungi on the growth of micropropagated potato plantlets and on the extent of diseases caused by Rhizoctonia solani. Mycorrhiza, 12, 235-242.
Pour citer cet article
A propos de : Badr Satrani
Centre de Recherche forestière. BP 763. Agdal. MA-10050 Rabat (Maroc). E-mail : badrsat@yahoo.fr
A propos de : Nouhad El Ouadihi
Centre de Recherche forestière. BP 763. Agdal. MA-10050 Rabat (Maroc) – Faculté des Sciences. Laboratoire de Botanique. MA-10040 Rabat (Maroc).
A propos de : Abdelhamid Guedira
Centre de Recherche forestière. BP 763. Agdal. MA-10050 Rabat (Maroc).
A propos de : Pascale Frey-Klett
Centre INRA de Nancy. Unité Mixte de Recherche Interactions Arbres/Micro-organismes (UMR IAM). F-54280 Champenoux (France).
A propos de : Mustapha Arahou
Faculté des Sciences. Laboratoire de Botanique. MA-10040 Rabat (Maroc).
A propos de : Jean Garbaye
Centre INRA de Nancy. Unité Mixte de Recherche Interactions Arbres/Micro-organismes (UMR IAM). F-54280 Champenoux (France).
