


- Home
- Volume 11 (2007)
- Numéro 1
- Evaluation de la diversité morphologique des variétés traditionnelles de sorgho du Nord-ouest du Maroc


View(s): 5921 (43 ULiège)
Download(s): 470 (9 ULiège)

Evaluation de la diversité morphologique des variétés traditionnelles de sorgho du Nord-ouest du Maroc

Editor's Notes
Reçu le 25 mai 2005, accepté le 28 septembre 2006.
Résumé
Cette étude porte sur la variabilité des types morphologiques de variétés traditionnelles de sorgho récoltées dans 5 champs sous culture traditionnelle au nord ouest du Maroc. Vingt-deux caractères qualitatifs et quantitatifs ont servi à évaluer la diversité de 125 plantes. Une importante variabilité morphologique est observée entre les champs. Les caractères ayant trait au rendement (DIAM, POIDS), à la croissance végétative (HAU, LOF, LAF, DIA) et à la panicule (LOP, PED) sont celles qui influencent le plus la variabilité observée. L’utilisation par les paysans de leurs semences propres pour la culture associée à la sélection paysanne basée sur des caractères phénotypiques joueraient un rôle important dans le maintien de la diversité entre champs. La pérennité du sorgho traditionnel dans une région marginale comme le nord ouest du Maroc nécessite la mise en place rapide de programme de conservation in situ et ex situ pour sauvegarder cette richesse génétique.
Abstract
Evaluating morphological diversity of sorghum landraces from North Western Morocco. In this study, we assessed the pattern of variation in spikelet morphology and vegetative characters of sorghum landraces sampled in situ from 5 fields under traditional cultivation system in North Western Morocco. We use 22 quantitative and qualitative morphological traits to evaluate diversity for an amount of 125 individual plants. Data showed a great morphological variability among fields due to yield parameters (DIAM, POIDS), growth characters (HAU, LOF, LAF, DIA) and spikelet morphology (LOP, PED). The use of their own seeds for culture combined with peasants’ selection based on phenotypic characters would play an important role in persistence of diversity among farmers’ fields. Since farmers from North Western Morocco use a marginal cropping system for sorghum, we suggest the implementation of an in situ and ex situ conservation program to protect this genetic richness.
Table of content
1. Introduction
1Les variétés traditionnelles des espèces cultivées sont en voie de disparition dans plusieurs régions du monde tropical. En effet, l’introduction des cultures de rente et de variétés améliorées à haut rendement a contribué à restreindre l’aire de culture des variétés traditionnelles qu’on retrouve uniquement dans les campagnes les plus reculées (Brush, 1995). Les études de diversité des plantes cultivées ont montré clairement que les variétés traditionnelles, bien que moins productives, sont génétiquement plus diversifiées que les variétés améliorées (Brown, Munday, 1982 ; Ahmadi et al., 1988) et leur sauvegarde contribue au maintien de cette richesse génétique (Frankel et al., 1995). Pour pallier cette disparition et éviter une catastrophe biologique, de nombreuses accessions de variétés traditionnelles ont été récoltées et conservées dans des banques de gènes (Harlan, 1975 ; Frankel et al., 1995). Ces accessions maintenues et reproduites ex situ ont servi de matériel biologique dans des études de diversité morphologique (Chantereau et al., 1989), enzymatique (Maquet et al., 1997) et moléculaire (Fofana et al., 1997). Cependant en raison de cette longue phase de conservation, la variabilité génétique observée ne reflète pas toujours la diversité présente in situ (Doebley et al., 1985 ; Djè et al., 1998). Il convient dans la mesure du possible de porter un regard nouveau sur le matériel végétal dans son aire naturele de culture surtout en ce qui concerne les espèces cultivées. Cela est urgent pour les études morphologiques car les reproductions en serres, même tropicalisées, n’offrent jamais les mêmes conditions que celles rencontrées dans les pays d’origine.
2Le sorgho (Sorghum bicolor L. Moench) est cultivé dans plusieurs régions tropicales du monde où il entre dans la consommation humaine, animale et dans les industries. En rapport avec la surface cultivée qui lui est consacrée, le sorgho est une céréale cultivée principalement sous les tropiques. Le sorgho cultivé revêt une importance capitale dans les zones sahéliennes d’Afrique où il représente l’une des principales ressources alimentaires. Son appareil racinaire important et profondément ancré dans le sol lui permet d’aller puiser l’eau souterraine pour assurer son bon développement dans un environnement souvent hostile (Doggett, 1988 ; Chantereau, Nicou, 1991). à l’échelle du Maroc, le sorgho n’est pas une culture céréalière importante. Il est cultivé et maintenu comme culture ancestrale uniquement dans le nord ouest du pays où il est utilisé dans l’alimentation humaine et animale. Le sorgho, appelé « draa » en langue locale, y est cultivé selon des pratiques culturales anciennes sans intervention d’intrants et d’outils agricoles modernes. Les paysans classent les sorghos locaux principalement en fonction de la couleur des grains et de la farine qui en est extraite. Ils distinguent ainsi les sorghos à farine rouge « draa hamra » et les sorghos à farine blanche « draa bayda ». Le type de panicule (lâche à compacte) constitue le second critère de distinction au sein de sorgho ayant la même couleur de grain. Les petits producteurs conservent une partie de leurs semences pour la culture de l’année suivante. Les échanges de semences restent exceptionnels et dans ces cas, le ravitaillement se fait chez le voisin immédiat ou à défaut sur le marché local. Les champs de sorgho morcelés et de taille modeste, sont situés sur le flanc des montagnes sur des sols ferralitiques ou dans les plaines sur des sols limoneux. Les semis sont réalisés avec les premières pluies du printemps et la récolte a lieu au cours de l’été (fin août à début septembre). Le sorgho est l’une des principales céréales dans le nord ouest du Maroc où il représente la principale céréale de printemps mais cependant très peu d’études ont concerné cette culture (Kadiri, Ater, 1997). Les études de diversité réalisées par Djè et al. (1998) portant sur 10 caractères qualitatifs et 54 champs ont montré l’existence d’une variabilité morphologique plus importante entre les champs que celles observées à partir des données enzymatiques. Le présent travail concerne une étude portant sur un échantillonnage plus large et analysé sur la base d’un nombre plus élevé de caractères qualitatifs et quantitatifs. Les résultats obtenus seront discutés en tenant compte de la nature des caractères analysés ainsi que de la spécificité du mode de conservation des semences destinées à la culture.
2. Matériels et méthodes
2.1. Matériel végétal et zone d’étude
3La figure 1 montre les localités où les échantillons ont été collectés. La zone de prospection couvre les latitudes 35 à 36 °N et les longitudes 5 à 6 °E et comprend les provinces de Tétouan et Chechaouen (Tableau 1). Les individus sont choisis dans les champs de façon aléatoire à partir d’un point puis les différentes mesures de caractères végétatifs sont notées in situ. Ensuite la panicule entière de l’individu cible est coupée pour les mesures réalisées au laboratoire. Au total nous avons analysé 25 individus par champ, soit 125 plantes.
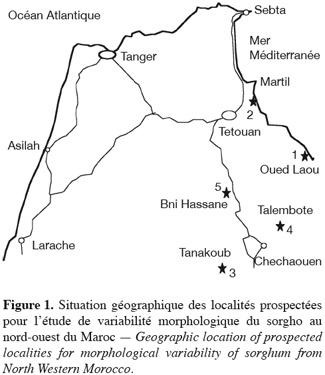
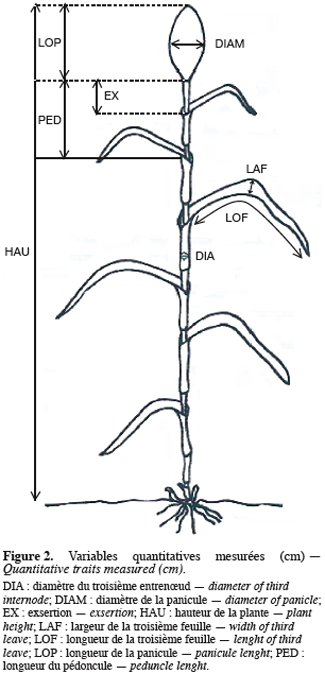
42.2. Méthodes de mesures des caractères
5Deux catégories de caractères ont été étudiées. Il s’agit de caractères quantitatifs et de caractères qualitatifs. La figure 2 présente la méthode de mesure de 8 des 11 caractères quantitatifs mesurés (le diamètre du troisième entrenœud en cm, DIA ; le diamètre de la panicule en cm, DIAM ; l’exsertion en cm, EX ; la hauteur de la plante en cm, HAU ; la largeur de la troisième feuille en cm, LAF ; la longueur de la troisième feuille en cm, LOF ; la longueur de la panicule en cm, LAP et la longueur du pédoncule en cm, PED). Le poids de la panicule en g (POIDS), le poids de 100 grains en g (P100) et le nombre d’entrenœuds (NEN) sont les 3 autres caractères mesurés. On distingue parmi ces caractères ceux purement végétatifs (HAU, LOF, LAF, DIA et NEN) et ceux concernant la panicule (LOP, PED, EX, DIAM, POIDS et P100). Parmi ceux-ci, les 3 derniers (DIAM, POIDS et P100) concernent parti-culièrement le rendement. Les caractères qualitatifs évalués sont également au nombre de 11 (Tableau 2) et concernent :
6– l’inflorescence : la forme du pédoncule (FPED) et le type de panicule (TPAN) ;
7– le grain : la forme (FGR), l’allongement (AGR), la couleur (CGR), le pourcentage de recouvrement par les glumes (REC) et le pourcentage de réserves amylacées de l’endosperme (END) ;
8– l’épillet : la taille de l’arête (ARE), la couleur (CGL), la pilosité (PIL) et l’aspect de la glume (AGL).
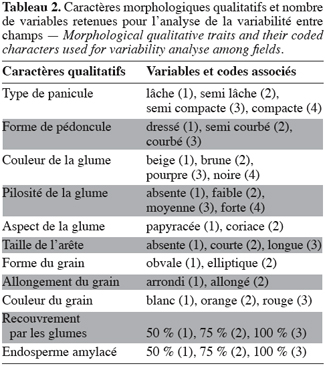
2.3. Analyses statistiques
9Pour chacun des caractères quantitatifs étudiés, nous avons procédé à une comparaison des moyennes par l’analyse de la variance (ANOVA). Lorsqu’une différence significative est révélée entre champs pour un caractère donné, l’ANOVA est complétée par le test de Newman-Keuls qui permet d’identifier le ou les champ(s) qui diffère(nt) significativement des autres. Le coefficient de variation permet d’apprécier les niveaux de variation des moyennes observées entre les champs pour chaque caractère. L’ensemble des 22 caractères a servi à réaliser une analyse en composantes principales (ACP). L’ACP permet, à partir de la corrélation des matrices des variables originelles, d’extraire un petit nombre de combinaisons linéaires non corrélées entre elles. Les composantes principales sont construites de manière à rendre compte de la plus grande fraction de la variance totale. On retient pour l’analyse les premières composantes principales qui prennent en compte la majeure partie de la variance observée. La détermination, pour chaque composante principale, des variables qui y sont fortement corrélées, permet de ressortir les variables originelles qui contribuent le plus à la valeur de chaque composante principale (Manly, 1994). La projection de l’ensemble des individus sur les plans des principaux axes des principales composantes permet ensuite d’apprécier la dispersion des individus et de mieux comparer la variabilité entre les champs.
3. Résultats
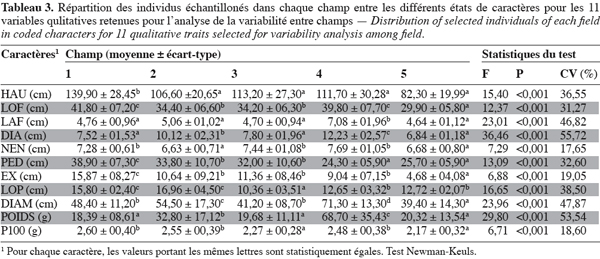
10L’analyse de la variance des caractères quantitatifs est regroupée dans le tableau 3. On observe pour chaque caractère une différence hautement significative entre les différents champs (P < 0,001) qui est la résultante d’une diversité morphologique entre les individus qui composent ces champs. Les valeurs du coefficient de variation indiquent une faible variation entre les champs pour le nombre d’entrenoeuds (NEN), le poids de 100 grains (P100) et l’exsertion (EX) (respectivement 17,65 %, 18,60 % et 19,05 %). à l’inverse, on observe une forte variation entre les champs pour le diamètre du troisième entrenoeud (DIA), le poids de la panicule (POIDS) et la largeur de la troisième feuille (LAF) (respectivement 55,72 %, 53,64 % et 48,62 %). Cela indique une très forte hétérogénéité entre les champs pour ces 3 caractères. Les individus du champ 5 ont les plus faibles valeurs végétatives et des panicules de faible diamètre constituées des grains les plus petits. Le champ 1 présente des individus de grande taille à caractères végétatifs très développés et les panicules les plus légères constituées des grains les plus gros. Ces caractéristiques se retrouvent dans le champ 2 où les individus ont cependant des panicules lourdes et larges. Les rendements les plus importants sont observés dans le champ 4 avec des individus à caractères végétatifs très développés. Les individus du champ 3 ont des petites panicules et un port végétatif moyen.
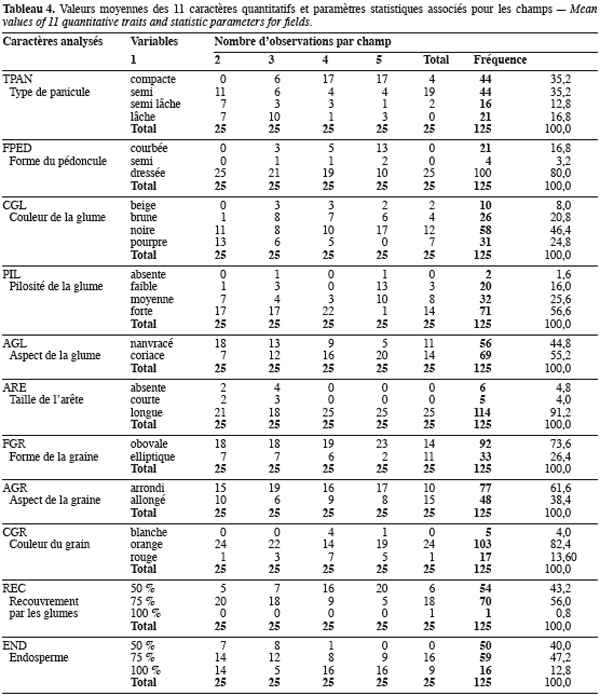
11La fréquence d’apparition des différents types morphologiques à l’intérieur de chaque caractère qualitatif au niveau de chaque champ et sur l’ensemble des individus échantillonnés est présentée dans le tableau 4. On observe que les caractères taille de l’arête (ARE), couleur du grain (CGR) et forme du pédoncule (FPED) sont les moins variables sur l’ensemble des 125 individus analysés. En effet, pour chacun de ces 3 caractères, on observe une prédominance d’un type morphologique donné. Ainsi, 91,2 % (114/125) des individus ont une arête longue (ARE), 82,4 % (103/125) possèdent des grains de couleur orange (CGR) et 80 % (100/125) ont un pédoncule dressé (FPED). La tendance à la prédominance d’un type morphologique donné, pour chacun de ces 3 caractères, est observée au niveau de chaque champ à l’exception notable de la forme du pédoncule (FPED) pour laquelle on observe une prédominance des panicules courbées et semi courbées (60 %) parmi les individus du champ 4. Les caractères les plus variables sont le type de panicule (TPAN), la couleur de la glume (CGL) et le type d’endosperme (END). Pour ces caractères, on n’observe pas une prédominance particulière d’un type morphologique parmi les individus analysés. Pour les autres caractères (la pilosité des glumes, PIL ; la forme de la graine, FGR ; l’aspect des glumes, AGL ; l’aspect des graines, AGR), on remarque qu’un type morphologique est assez bien représenté parmi les individus mais cela n’empêche pas la présence significative des autres types au niveau de chaque caractère. L’analyse de la fréquence d’observation de chaque type morphologique constituant les différents caractères qualitatifs au niveau des champs permet de dégager 2 groupes morphologiques de champs. Le premier groupe est constitué des champs 3 et 4 caractérisés par une prédominance des panicules compactes à glume d’aspect coriace recouvrant seulement la moitié des grains dont les réserves sont entièrement amylacées. Le second groupe est constitué par les champs 1, 2 et 5 caractérisés par des individus à panicules lâches, à glumes d’aspect papyracé qui recouvrent presque la totalité des grains dont les réserves sont partiellement cornées.
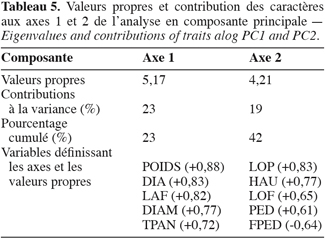
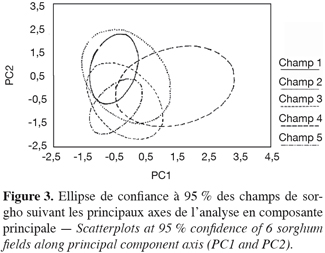
12L’analyse en composantes principales de l’ensemble des caractères morphologiques mesurés révèle que les 4 premières composantes regroupent 58 % de l’inertie totale (respectivement 23 % pour la première composante principale, PC1 ; 19 % pour PC2, 9 % pour PC3 et 7 % pour PC4). Les caractères fortement corrélés au premier axe (r > 0,6) sont le poids de la panicule (POIDS), le diamètre du troisième entrenoeud (DIA), la largeur de la troisième feuille (LAF), le diamètre de la panicule (DIAM) et le type de panicule (TPAN). Le deuxième axe est fortement corrélé positivement à la longueur de la panicule (LOP), la hauteur de la plante (HAU), la longueur de la troisième feuille (LOF), la longueur du pédoncule (PED) et corrélé négativement à la forme du pédoncule (FPED) (Tableau 5). Les caractères fortement corrélés à l’axe 1 ont trait à des aspects végétatifs (TPAN, LAF et DIA) ou au rendement (POIDS et DIAM). Les variables fortement corrélées positivement à l’axe 2 sont toutes liées à la taille de la plante (LOP, HAU, LOF et PED). La figure 3 montre la projection des ellipses de confiance à 95 % de tous les individus des 5 champs sur le plan des 2 premières composantes principales (PC1 et PC2). On observe que les individus occupent toute la surface du plan. Les individus du champ 5 sont regroupés du côté négatif des 2 axes où on retrouve peu d’individus du champ 4. Ceux-ci sont largement éparpillés du côté positif de l’axe 1. Cette dispersion par rapport aux principaux axes (PC1 et PC2) des individus de ces 2 champs confirme les résultats de l’ANOVA suggérant que les champs 4 et 5 sont constitués d’individus présentant les plus grandes différences morphologiques. Le champ 5 est constitué d’individus grêles à faible rendement, ce qui l’oppose au champ 4 dominé par des individus à port végétatif développé avec un rendement élevé. Le champ 1 occupe une partie assez restreinte du plan et se situe entièrement dans la partie positive de l’axe 2, ce qui correspond à des individus à forte croissance végétative. L’aire de dispersion du champ 1 est assez restreinte et est entièrement comprise dans celui du champ 2. Les individus des champs 2 et 3 sont les mieux dispersés sur les 2 axes (PC1 et PC2) avec une tendance vers les valeurs négatives pour le champ 3 et les valeurs positives pour le champ 2. Les individus du champ 5 sont presque tous inclus dans l’aire de dispersion du champ 3, ce qui dénote d’une ressemblance entre les individus des 2 champs. Les individus du champ 4 sont les seuls à être dispersés dans presque leur totalité du côté positif de l’axe 1.
4. Discussion
13Les caractéristiques végétatives et paniculaires (caractères quantitatifs) des champs peuvent être mises en relation avec la localisation, la topographie et la fertilité des sols. Ainsi, il apparaît que les champs 1 et 2, situés dans les plaines fertiles de basse altitude en bordure de mer sont caractérisés par un développement végétatif important ou des caractéristiques paniculaires importantes. à l’inverse, les champs 3 et 5, situés sur des sols superficiels de la région de moyenne montagne du Rif, se caractérisent par des caractéristiques paniculaires faibles et un aspect végétatif peu développé. Le champ 4, situé dans la plaine alluviale riche en limon de l’intérieur du pays a les caractéristiques végétatives et paniculaires les plus avantageuses et apparaît intermédiaire entre ces 2 groupes. Les caractères quantitatifs sont ainsi étroitement liés aux conditions écologiques des zones de culture. à l’inverse, aucune relation particulière n’est observée entre les caractères qualitatifs étudiés et les conditions agroécologiques des sites de culture. Ainsi, les champs 1 et 2 situés sur des sols riches de la plaine ont les mêmes caractéristiques que le champ 5 situé sur un sol superficiel en pente. Les caractères morphologiques fortement corrélés (r > 0,6) aux axes des composantes principales 1 et 2 (PC1 et PC2) sont dominés par les caractères quantitatifs (8 caractères quantitatifs contre 2 caractères qualitatifs). Ceci pourrait résulter de la sélection phénotypique paysanne qui prend en compte principalement les caractéristiques végétatives et paniculaires. Ces caractères sont les plus remarquables au niveau des champs et semblent influencer grandement les choix des paysans. En effet, selon l’utilisation qui en sera faite, les sorghos présentant les caractéristiques paniculaires les plus avantageuses ou les caractéristiques végétatives les plus importantes seront ceux dont les grains seront préférentiellement conservés pour les semis à venir. Les premiers seront préférés par ceux qui destinent leurs récoltes à la consommation humaine et les seconds par ceux qui utilisent leurs cultures comme fourrage. Les caractères quantitatifs, bien que soumis à l’influence des conditions environnementales, ne doivent pas être négligés par les conservateurs dans l’étude de la diversité car ils sont essentiels en milieu paysan où ils influencent grandement les critères de sélection phénotypique.
14Il est intéressant de constater que les seuls caractères qualitatifs qui influent significativement sur les axes de la première et de la deuxième composante principale (PC1 et PC2) sont le type de panicule (TPAN) et la forme du pédoncule (FPED). En effet, parmi les 11 caractères qualitatifs étudiés et à l’exception de la couleur du grain (CGR), ces 2 caractères sont ceux qui sont phénotypiquement les plus perceptibles par les paysans au niveau de leurs champs. Ainsi, des caractères comme le type d’endosperme (END) ou l’aspect de la glume (AGL) ne sont pas pris en compte dans la sélection phénotypique paysanne. Le type de panicule (lâche à compacte) et la forme du pédoncule (dressée ou courbée) sont à l’inverse perceptibles dans les champs et peuvent ainsi orienter la sélection phénotypique paysanne. Le caractère type de panicule (TPAN), non directement influencé par les conditions du milieu et perçu par les paysans eux-mêmes comme second critère de distinction des pieds de sorgho, peut servir valablement à étudier la variabilité des sorghos du Maroc. En effet, il existe un large éventail de types morphologiques associées aux types de panicule à l’intérieur des sorghos cultivés allant du type lâche « balai de sorcière » au type compact (Harlan, de Wet, 1972).
15Les pourcentages de variance exprimés par les 4 principaux axes de l’analyse en composante principale (23 à 7 %) indiquent l’absence d’un caractère morphologique prépondérant dans la variabilité des sorghos du nord ouest du Maroc. à partir des études morphologiques de 10 caractères de l’inflorescence et portant sur 54 panicules récoltées dans 6 champs, Djè et al. (1998) ont montré que, malgré l’existence d’une importante variabilité au sein des sorghos locaux, les pourcentages de variance exprimés par les principaux axes (PC1 à PC4) étaient assez faibles et compris entre 22 et 11 %. Selon Idrissi et Ouazzani (2003), une faible représentativité des axes témoigne de l’absence d’une organisation génotypique et phénotypique forte entre les champs étudiés. Cette absence d’organisation forte peut s’expliquer par l’origine monophylétique des sorghos du nord ouest du Maroc. L’hypothèse la plus probable actuellement admise est celle liée à l’invasion arabe. En effet, la classification actuelle des sorghos cultivés distingue 5 races avec une série d’hybrides entre ces races (Harlan, de Wet, 1972). La répartition des types actuels de sorgho montre que le type principalement cultivé au Moyen Orient correspond à la race durra (Doggett, 1988). Les études de Kadiri et Ater (1997) et Djè et al. (1998) ont montré l’appartenance des types observés au nord ouest du Maroc à la race durra et à des hybrides inter races incluant la race durra. Les études préliminaires de Kadiri et Ater (1997) à partir de 431 individus issus de 27 champs et analysés par 11 caractères de l’inflorescence et de l’épillet ont montré l’existence d’une importante variabilité au sein des sorghos traditionnels du Maroc qui sont constitués de 2 cultivars de la race durra dont les aires de culture peuvent se superposer avec la répartition des populations de la région nord ouest du Maroc. Le cultivar durra est cultivé par des populations arabes et le cultivar cernuum par des tribus composites. Une telle concordance entre les zones de culture et les types traditionnels de sorgho cultivé a été observée par Teshome et al. (1997) en Ethiopie. Ils ont observé une très grande diversité in situ entre les types morphologiques cultivés et une consistance entre les différents noms vernaculaires locaux et les grandes races de sorgho cultivé qui résulte d’une longue tradition ancestrale.
16La présence d’une importante variabilité morphologique entre les champs peut être mise en relation avec le mode de conservation des semences. En effet, l’absence quasi-totale d’échanges de semences entre les différentes régions de culture favorise le maintien d’une variabilité entre les champs. Chaque population évolue de façon isolée des autres, ce qui accentue les différences observées au fil des cultures. Cette situation est accentuée par le régime de reproduction des sorghos du nord ouest du Maroc qui sont préférentiellement autogames avec un faible taux d’allogamie (10-20 %). On y observe aussi une structuration des champs en sous-populations qui se caractérisent par des niveaux d’allogamie très variables, des plantes entièrement autogames coexistant avec des plantes admettant un pourcentage très significatif d’allogamie (Djè et al., 2004). Les échanges de gènes entre champs voire à l’intérieur des champs restent ainsi limités, ce qui, combiné à la sélection paysanne accentuerait la particularité de chacun d’entre eux.
5. Conclusions et perspectives
17Une diversité considérable de types morphologiques est observée entre les champs de sorgho traditionnel du nord ouest du Maroc. La variabilité observée entre les champs étudiés constitue un argument de choix pour la conservation des variétés traditionnelles de sorgho du nord ouest du Maroc. Cette variabilité, dans une région où la culture du sorgho régresse d’année en année fait ressortir l’urgence de la mise en place de stratégies pour encourager la conservation in situ en milieu paysan. La position centrale des variétés locales dans les ressources végétales des plantes cultivées n’étant plus à démontrer (Frankel et al., 1995), une attention particulière doit être accordée aux régions qui assurent la pérennité de cette importance source de variabilité pour éviter une catastrophe scientifique.
Remerciements
18La réalisation de cette étude a été rendue possible grâce à un financement conjoint de l’Université Libre de Bruxelles (Belgique) et l’Université Abdelmalek Essaàdi de Tétouan (Maroc) dans le cadre d’une collaboration scientifique bilatérale.
Bibliographie
Ahmadi N., Becquer T., Larroque C., Arnaud M. (1988). Variabilité génétique du riz (Oryza sativa L.) à Madagascar. Agron. Trop. 43, p. 209–221.
Brown AHD., Munday J. (1982). Population-genetic structure and optimal sampling of land races of barley from Iran. Genetica 58, p. 85–96.
Brush SB. (1995). In situ conservation of landraces in centers of diversity. Crop Sci. 35, p. 346–354.
Chantereau J., Nicou R. (1991). Le sorgho. Paris : Maisonneuve et Larose, 160 p.
Chantereau J., Arnaud M., Ollitrault P., Nabayaogo P., Noyer JL. (1989). étude de la diversité morphophysiologique et classification des sorghos cultivés. Agron. Trop. 44, p. 223–232.
Djè Y., Ater M., Lefèbvre C., Vekemans X. (1998). Patterns of morphological and allozyme variation in sorghum landrace of northwestern Morocco. Genet. Res. Crop. Evol. 45, p. 541–548.
Djè Y., Heuertz M., Ater M., Lefèbvre C., Vekemans X. (2004). In situ estimation of outcrossing rate in sorghum landraces using microsatellite markers. Euphytica 138, p. 205–212.
Doebley J., Goodman M., Stuber C. (1985). Isozyme variation in the races of maize from Mexico. Amer. J. Bot. 75, p. 629–639.
Doggett H. (1988). Sorghum (2nd ed.). London: Longman Scientific and Technical.
Fofana B., Vekemans X., du Jardin P., Baudoin JP. (1997). Genetic diversity in Lima bean (Phaseolus lunatus L.) as revealed by RAPD markers. Euphytica 95, p. 157–165.
Frankel O., Brown AHD., Burdon JJ. (1995). The conservation of plant biodiversity. New York, USA: Cambridge University Press, 299 p.
Harlan JR. (1975). Our vanishing genetic resources. Science 188, p. 618–621.
Harlan JR., de Wet JMJ. (1972). A simplified classification of cultivated sorghum. Crop Sci. 12, p. 172–176.
Idrissi A., Ouazzani N. (2003). Apport des descripteurs morphologiques à l’inventaire et à l’identification des variétés d’olivier (Olea europea L.). Plant Genet. Resour. Newsl. 136, p. 1–10.
Kadiri M., Ater M. (1997). Diversité des variétés « locales » du sorgho grain (Sorghum bicolor Moench L.) au nord ouest du Maroc. In Rejdal M., Birouk H. (eds). Ressources phytogénétiques et développement durable. Actes Rabat, Maroc, p. 203–218.
Manly B. (1994). Multivariate statistical methods. London: Chapman & Hall,
Maquet A., Zoro Bi I., Delvaux M., Wathelet B., Baudoin JP. (1997). Genetic structure of a Lima bean base collection using allozyme markers. Theor. Appl. Genet. 95, p. 980–991.
Teshome A., Baum BR., Fahrig L., Torrance JK., Arnason TJ., Lambert JD. (1997). Sorghum (Sorghum bicolor (L.) Moench) landrace variation and classification in north Shewa and south Welo, Ethiopia. Euphytica 97, p. 255–263.
To cite this article
About: Yao Djè
Université d’Abobo-Adjamé. Unité de Formation et de Recherche des Sciences de la Nature.
02 BP 801 Abidjan 02 (Côte d’Ivoire). E-mail : saticlan@yahoo.fr
About: Myriam Heuertz
Laboratoire de Génétique et écologie végétales. Université libre de Bruxelles. 1850, chaussée de Wavre. B-1160 Bruxelles (Belgique).
About: Mohamed Ater
Université Abdelmalek Essaâdi. Faculté des Sciences. Département de Biologie. BP 2062, Tétouan 93002 (Maroc).
About: Claude Lefebvre
Laboratoire de Génétique et écologie végétales. Université libre de Bruxelles. 1850, chaussée de Wavre. B-1160 Bruxelles (Belgique).
About: Xavier Vekemans
Laboratoire de Génétique et écologie végétales. Université libre de Bruxelles. 1850, chaussée de Wavre. B-1160 Bruxelles (Belgique).
