


- Accueil
- volume 12 (2008)
- numéro 4
- La nécrose apicale en relation avec certains paramètres morphologiques et la teneur en calcium dans les fruits chez quatre variétés de piment (Capsicum annuum L.)


Visualisation(s): 3996 (27 ULiège)
Téléchargement(s): 222 (2 ULiège)

La nécrose apicale en relation avec certains paramètres morphologiques et la teneur en calcium dans les fruits chez quatre variétés de piment (Capsicum annuum L.)

Notes de la rédaction
Reçu le 30 août 2007, accepté le 30 janvier 2008
Résumé
L’objectif de cette étude est d’établir une relation entre la croissance végétative, le poids et la teneur en calcium des fruits et la sensibilité à la nécrose apicale chez quatre variétés de piment développant des fruits de poids unitaires différents : ' J27 ', ' Marconi ', ' Beldi ' et ' Jerid '. Les résultats obtenus montrent que les variétés les plus sensibles sont ' Marconi ' et ' J27 '. Celles-ci ont d’une part un poids, une vitesse de croissance du fruit et un taux de transpiration plus importants que les variétés à petits fruits et à vitesse de croissance plus faible, cas du ' Jerid '. D’autre part, leur teneur en calcium, surtout dans la partie apicale, est plus faible que chez les variétés à petits fruits, comme le ' Jerid '.
Abstract
Blossom-end rot in relation to morphological parameters and calcium content in fruits of four pepper varieties (Capsicum annuum L.). The objective of this study is to identify, in the case of four pepper cultivars: ' J27 ', ' Marconi ', ' Beldi ' and ' Jerid ', a relation between vegetative growth rate, fruit weight and its calcium content with blossom-end rot induction. The results show that the more sensitive cultivars to blossom-end rot are ' Marconi ' and ' J27 ' cultivars, those having a larger final size and faster growth speed of the fruits and a higher rate of transpiration in comparison with ' Jerid ' cultivar, which is characterized by its small fruits. On the other hand, the calcium contents of their fruit, especially in the apical part, is weaker than the ' Jerid ' cultivar’s one.
Table des matières
1. Introduction
1La nécrose apicale ou Blossom-end rot (BER) chez la tomate et le piment (Morley et al., 1993) est le symptôme d’un désordre physiologique lié à une déficience locale en calcium dans la partie apicale du fruit (Marcelis et al., 1999). Ce déficit est dû à un faible approvisionnement ou à un transport réduit de cet élément au niveau de cette partie du fruit, même quand la teneur en calcium dans la plante est élevée (Bradfield et al., 1984 ; Ho et al., 1995 ; Roca et al., 1997). Nonami et al. (1995) ont montré que la faible teneur en calcium dans cette partie n’est pas la seule cause de l’apparition de ce désordre. D’autres facteurs, indépendamment de la teneur en calcium dans la plante, peuvent induire la nécrose apicale (Ho et al., 1993) tels que la lumière, la température, l’humidité de l’air (R’him et al., 2005), l'humidité du sol (R’him et al., 2002), la taille et la vitesse de croissance du fruit (Marcelis et al., 1999). Tous ces facteurs peuvent favoriser l’apparition de la nécrose chez plusieurs espèces, entre autres le poivron. En effet, plusieurs auteurs (Mix et al., 1976 ; Tromp, 1979) ont observé une corrélation négative entre les dimensions des fruits et la concentration en calcium. Cette corrélation négative peut être expliquée par une augmentation du transport des assimilats par la voie phloémique sans augmentation du transport de Ca par la voie xylémique pendant la phase de croissance rapide du fruit (Marcelis et al., 1999). A ce stade critique, il y a une plus grande demande en calcium pour la croissance et la multiplication rapide des cellules excédant l’approvisionnement en Ca nécessaire aux tissus des fruits (Wiersum, 1966 ; Ho, 1980 ; Ehret et al., 1986 ; Ho et al., 1993 ; Ho et al., 2005). Ho et al. (1987) ont signalé pour la tomate que les gains de matière sèche et d’eau dans le fruit sont principalement assurés par le phloème alors que le transport de Ca est limité au xylème. Par conséquent, un déséquilibre entre les assimilats des feuilles et le Ca fourni a été la cause commune de l’induction de la nécrose apicale chez le poivron et la tomate (Marcelis et al., 1999). Roca (1996) et Roca et al. (1997) ont montré que le facteur variétal reste aussi le plus important élément risquant d’occasionner la nécrose apicale dans les cultures, particulièrement chez le poivron. Ils ont expliqué que les variétés dites sensibles auraient tendance à avoir un contenu plus faible en calcium dans le fruit (Sanchez Conde et al., 1983 ; Franco et al., 1994 ; Cardoso et al., 1995 ; Willumsen et al., 1996) ; une vigueur végétative importante et des fruits de gros calibres (Ho et al., 1993 ; Morley et al., 1993 ; Marcelis et al., 1999). En effet, Robbins (1937) et Saure (2001) ont observé que les fruits pygmés (fruits à petits calibres dont le poids et la longueur ne dépassent pas respectivement 10 g et 5 cm) n’ont aucune phase de croissance rapide et ne développent pas de nécrose apicale. De même, les variétés vigoureuses peuvent réduire l’importation de Ca dans les fruits et par la suite la fraction de Ca transportée aux feuilles (concurrence inter-puits au profit des organes transpirants) menant à une plus grande sensibilité à la nécrose apicale (Aikman et al., 1990).
2L’objectif de cette étude est d’évaluer l'importance relative de la nécrose apicale chez quatre variétés de piment en relation avec la transpiration, la surface foliaire, le poids sec final de la plante d’une part, la vitesse de croissance, le poids et le contenu en calcium dans le fruit d’autre part.
2. Matériel et méthodes
2.1. Matériel végétal
3Le matériel végétal utilisé dans ce travail est constitué de quatre variétés de piment présentant des fruits de tailles différentes appartenant à la forme cultivée Capsicum annuum. Deux variétés locales : ' Beldi ' (longueur moyenne du fruit = 15 cm, poids moyen du fruit = 23 g) et ' Jerid ' (longueur moyenne du fruit = 4 cm, poids moyen du fruit = 4 g) cultivées respectivement en culture de saison en plein champ et en été au Sahel (Centre-Est de la Tunisie) et au sud tunisien. L’hybride F1 : ' J27 ' (longueur moyenne du fruit = 20 cm, poids moyen du fruit = 35 g) créé par l’INRAT (Tunisie) et le poivron de type long, ' Marconi ' (longueur moyenne du fruit = 20 cm, poids moyen du fruit = 65 g), d’origine italienne, sont cultivés essentiellement sous abri-serres plastiques.
2.2. Conduite de la culture
4L’expérimentation est réalisée dans une serre vitrée à la station expérimentale de Mornag (Nord-Est de Tunis) relevant de l’Institut National de la Recherche Agronomique de Tunisie durant l’année 2003 et 2004.
5Le semis des quatre variétés est effectué au début du mois d’octobre en terrines remplies de tourbe, à 25 °C. Il est suivi, à environ 20 jours, d’un repiquage en plaques alvéolaires. La culture dure 30 jours. Elle est conduite à 20-21 °C de nuit et 28 °C de jour, sous une HR de 70 %. Au stade 4 feuilles (20 novembre), les plantes sont transférées dans des pots en plastique de 30 cm de diamètre et de 50 cm de profondeur, contenant chacun 13,5 kg de tourbe. Les plantes sont irriguées avec une solution fertilisante (en mM.l-1) contenant selon Martinez et al. (2001) : 9,9 NO-3 ; 1,5 H2PO4- ; 4,5 SO4- ; 0,9 NH4+ ; 7,5 K+ ; 4,5 Ca2+ ; 1,5 Mg2+ et 0,5 g.l-1 d’un complexe d’oligo-éléments.
6Durant toute la période de l’essai (du stade plantation au mois de novembre jusqu’à la fin de la culture au mois de mars), le substrat de culture est maintenu à la capacité au champ par des irrigations fréquentes afin d’éviter que l’eau ne soit un facteur limitant.
7Les pots sont placés dans une serre vitrée équipée d’un pyranomètre mesurant le rayonnement global solaire naturel (2500-3000 joule.cm-²) pendant les mois de culture ; d’un thermohygrographe de type Testo (Testostor 175) permettant la mesure de la température ambiante et de l’humidité relative de l’air (HR).
8Durant toute l’expérimentation, la température minimale de l’air a été maintenue à 28 °C pendant le jour (16 h) et 15° C pendant la nuit. L’humidité relative minimale de l’air a été maintenue à 60 % pendant le jour et 80 % pendant la nuit.
9Les fruits sont récoltés à l’état mûr et vert (maturité de consommation), lorsque le pédoncule se détache facilement de la plante. Trois récoltes à intervalle de 20 jours sont effectuées (1er février, 20 février et 10 mars).
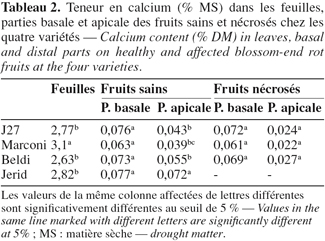
2.3. Dispositif expérimental
10Le dispositif expérimental est un essai factoriel en randomisation totale à 5 répétitions. Le premier facteur est défini par les quatre variétés de piment. Chaque répétition comprend trois pots par variété.
2.4. Les paramètres mesurés
11– La transpiration des plantes a été mesurée par un système portable de photosynthèse, modèle " CI-301 CO2 Gaz analyzer " (CID Inc., Camas, WA, USA), tout au long du cycle de la culture. Une description détaillée de l’installation a été faite par Huygens et al. (1992). La valeur moyenne calculée par variété correspond à trois mesures effectuées sur des feuilles appartenant aux deux derniers étages foliaires.
12– La surface foliaire et la matière sèche de la plante ont été déterminées à la fin de la culture par un planimètre électronique de type area Meter CI-202.
13– La matière sèche a été déterminée par pesée à l’aide d’une balance de précision de type Mettler P3 après séchage des échantillons à l’étuve à 80 °C pendant 48 heures.
14– Le rendement en fruits a été déterminé à chaque récolte en grammes par plante.
15– Le poids moyen du fruit a été calculé selon la formule suivante à chaque récolte :

16– Le poids sec du fruit a été déterminé après séchage des échantillons à l’étuve pendant 48 heures à la température de 80 °C.
17– La vitesse de croissance du fruit est estimée durant l’essai sur 15 échantillons pour chaque variété. Les mesures ont été faites 25 et 45 jours à partir de la nouaison des fruits (fruit noué lorsque le φ = 2 mm).
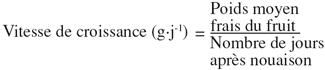
18– Le pourcentage de nécrose apicale est défini par le pourcentage des fruits non commercialisables. A chaque récolte, un premier triage est effectué sur les fruits présentant la nécrose apicale pour chaque variété. Leur importance est exprimée en pourcentage par rapport au nombre total des fruits récoltés.
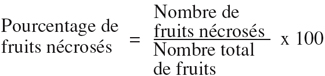
19– La teneur en calcium est déterminée par spectrophotomètre à flamme (Eppendorf).
20Le matériel végétal utilisé est constitué des fruits sains, fruits nécrosés et feuilles prélevées des plantes des différentes variétés (' J27 ', ' Marconi ', ' Jerid ' et ' Beldi ').
21Avant séchage des échantillons à 80 °C pendant 48 h à l’étuve, on procède à la division des fruits (sains et nécrosés) en deux parties : une zone pédonculaire et une zone pistillaire. Les échantillons séchés (feuilles, parties basale et apicale) sont mis dans des piluliers contenant 25 ml d’acide nitrique 0,1 N. L’extraction du calcium a lieu à la température ambiante du laboratoire pendant au moins 48 h. Les mesures sont exprimées en % de matière sèche.
2.5. Etude statistique
22L’analyse de la variance ANOVA a été faite par le programme STAT-ITCF. Chaque donnée constitue la moyenne d’au moins trois mesures. Pour la comparaison des moyennes, nous avons utilisé le test de Newman et Keuls au seuil de 5 %.
3. Résultats et discussion
3.1. Croissance végétative et transpiration
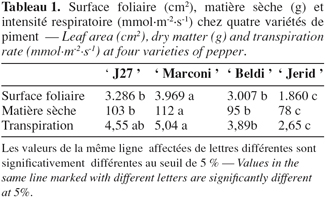
23L’analyse de la variance de la surface foliaire, de la matière sèche totale et de la transpiration montre une différence significative entre les variétés. La variété ' Marconi ' présente la surface foliaire, la matière sèche et le taux de transpiration les plus importants. Aucune différence significative pour ces paramètres entre ' J27 ' et ' Beldi '. Les valeurs les plus faibles sont obtenues chez la variété ' Jerid ' (Tableau 1).
3.2. Vitesse de croissance des fruits
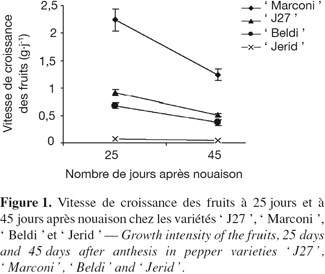
24L’observation de la figure 1 montre que la vitesse de croissance des fruits durant les 25 premiers jours juste après nouaison est supérieure de 45 % par rapport à celle de la deuxième période. La vitesse de croissance des fruits de la variété ' Marconi ' pendant les deux périodes est respectivement plus élevée de 2 fois et demi, 3 fois et demi et de 28 fois par rapport à celle de ' J27 ', de ' Beldi ' et de ' Jerid '.
3.3. Relation entre la sensibilité variétale à la nécrose apicale, la teneur en calcium et le poids du fruit
25En ce qui concerne la sensibilité variétale à la nécrose apicale, on constate (Figure 2A) que la variété ' Marconi ' qui donne les plus gros fruits (poids moyen/fruit > 60 g) (Figure 2B) est la plus sensible à ce désordre physiologique : le pourcentage de fruits nécrosés a dépassé 25 %. En revanche, la variété ' Jerid ' suivie de la variété ' Beldi ' dont les poids moyens par fruit ne dépassent pas respectivement 5 g et 25 g se sont montrées les plus résistantes à ce désordre physiologique. En effet, le pourcentage de fruits nécrosés est nul pour la variété ' Jerid ' et seulement de 4 % pour ' Beldi '.
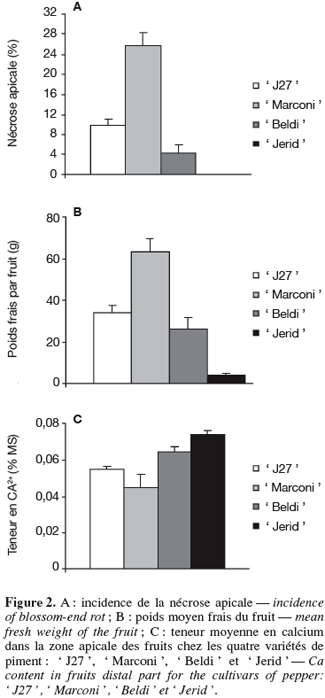
26Pour ce qui est de la teneur en calcium dans les fruits (Figure 2C), on remarque une corrélation négative entre la teneur en calcium et la sensibilité variétale à la nécrose apicale. En effet, on constate que la teneur la plus élevée de calcium est enregistrée chez ' Jerid ' (0,074 %) suivi de ' Beldi ' (0,064 %) et de ' J27 ' (0,055 %). La teneur la plus faible en Ca est notée chez les fruits de ' Marconi ' (0,045 %).
3.4. Comparaison de la teneur en calcium dans les feuilles, les parties basale et apicale des fruits
27L’observation du tableau 2 montre que les feuilles sont beaucoup plus riches en calcium que les fruits, surtout chez la variété ' Marconi '. Ceci est lié à l’origine de l’organe. Les feuilles sont les organes les plus transpirants, détournant ainsi le plus de calcium vers elles aux dépens des organes les moins transpirants, soit les fruits.
28Dans la partie pédonculaire des fruits sains et nécrosés, aucune différence variétale n’a été constatée dans la distribution du calcium.
29Le dosage du calcium dans la partie pistillaire des fruits sains et nécrosés montre que ce dernier est deux fois plus élevé dans les fruits sains que dans les fruits nécrosés. La teneur en calcium dans la partie apicale des fruits nécrosés n’est pas statistiquement différente chez les trois variétés, la teneur en calcium dans cette partie présentant la nécrose varie entre 0,022 et 0,027 %.
30En comparant la teneur en calcium trouvée dans les parties pédonculaire et pistillaire des fruits des quatre variétés, on constate qu’il y a des différences significatives entre ces deux parties chez les variétés à l’exception de ' Jerid '. En effet, la teneur en calcium de la partie basale des fruits de ' Marconi ', ' J27 ' et ' Beldi ' est plus élevée respectivement de 43, 39 et 25 % par rapport à la partie apicale.
4. Discussion
31Au cours de cette expérimentation, nous avons essayé de répondre à la question : existe-t-il une relation entre la croissance des plantes et le poids des fruits sur la prolifération de la nécrose apicale et la teneur en calcium chez quatre variétés de piment ?
32Trois résultats ressortent de ce travail.
33Le premier est que la sensibilité variétale à la nécrose apicale qui se manifeste dès deux semaines après l’anthèse (Manishi et al., 1996 ; Wada et al., 1996 ; Cho et al., 1998 ; Marcelis et al., 1999 ; Adams, 2002) est fonction de la vitesse de croissance et du poids des fruits. En effet, les résultats obtenus nous ont montré que la vitesse maximale de croissance des fruits et par suite le maximum de gain de biomasse des fruits se fait pendant les 25 premiers jours après nouaison, période qui coïncide avec l’apparition de la nécrose. Cette croissance rapide des fruits, principalement pour ' Marconi ', peut être expliquée par une augmentation du transport des assimilats par la voie phloémique suite à la transpiration plus importante de cette variété sans augmentation de transport de Ca par la voie xylémique pendant la phase de croissance des fruits (au cours des deux périodes). La demande en Ca pour l’agrandissement des cellules est augmentée (Ho et al., 1993 ; Morley et al., 1993 ; Marcelis et al., 1999 ; Ho et al., 2005) par un approvisionnement plus élevé en assimilats, alors que la quantité de calcium fournie au fruit n’augmente pas toujours proportionnellement. Ceci corrobore les observations de Marcelis et al. (1999) et Morley (1996) sur le poivron et de Adams et al. (1995), Ho et al. (1987) et Ehret et al. (1986) sur la tomate. Par contre, pour les variétés peu vigoureuses, telles que ' Beldi ' (fruits de poids moyen) ou ' Jerid ' (petites feuilles et fruits pygmés), leur développement ne passe pas par une phase rapide de croissance (Figures 1 et 2). La quantité de calcium arrivée aux fruits satisfait les besoins des fruits en cet élément. Ceci est en accord avec les résultats de Robbins (1937) et Saure (2001).
34Le deuxième résultat est que pour les variétés sensibles à la nécrose, chez qui les feuilles sont d’importants drains de calcium (Tableau 2), un taux élevé de transpiration (cas de ' Marconi ') augmente probablement, comme l’avaient indiqué plusieurs auteurs (Ho, 1989 ; Aikman et al., 1990 ; Paiva et al., 1998 ; Saure, 2001), la concurrence pour Ca2+ entre les feuilles et le fruit au profit des organes transpirants. Il distrait ainsi le flux transportant le Ca2+ du fruit et des jeunes pousses aux feuilles et mène à une plus grande sensibilité à la nécrose apicale.
35Le troisième résultat est que les variétés qui sont sensibles ont un contenu plus faible en calcium dans le fruit, surtout dans la partie apicale. Ceci est en accord avec les observations de Sanchez Conde et al. (1983), Franco et al. (1994), Cardoso et al. (1995) et Willumsen et al. (1996).
5. Conclusion
36A l’issue de cette étude, on peut dire que la sensibilité variétale à la nécrose apicale est due à plusieurs facteurs combinés :
37– une vitesse de croissance élevée des fruits, surtout pendant les trois semaines juste après nouaison,
38– une vigueur végétative importante et des fruits de gros calibre,
39– un taux élevé de transpiration,
40– un contenu plus faible en calcium dans la partie apicale du fruit.
Bibliographie
Adams P., 2002. Nutritional control in hydroponics. In: Savvas D. & Passam H., eds. Hydroponic production of vegetables and ornamentals. Athens: Embryo Publications, 211-261.
Adams P. & Ho L.C., 1995. Uptake and distribution of nutrients in relation to tomato fruit quality. Acta Hortic., 412, 374-387.
Aikman D.P. & Houter G., 1990. Influence of radiation and humidity on transpiration: implications for calcium levels in tomato leaves. J. Hortic. Sci., 65, 245-253.
Bradfield E.G. & Guttridge C.G., 1984. Effects of night-time humidity and nutrient solution concentration on the calcium content of tomato fruit. Sci. Hortic., 22, 207-217.
Cardoso M.O., de Queiroz M.A. & Souza R.F., 1995. Incidência de podridao apical em cinco cultivares de tomateiro cultivados em solo com três niveis de calcio. Hortic. Bras., 13, 172-175.
Cho I.H. et al., 1998. Effects of high humidity on occurrence of tomato blossom-end rot. J. Korean Soc. Hortic. Sci., 39, 247-249.
Ehret D.L. & Ho L.C., 1986. Translocation of calcium in relation to tomato fruit growth. Ann. Bot., 58, 679-688.
Franco J.A., Bañón S. & Madrid R., 1994. Effects of a protein hydrolysate applied by fertigation on the effectiveness of calcium as a corrector of blossom-end rot in tomato cultivated under saline conditions. Sci. Hortic., 57, 283-292.
Ho L.C., 1980. Control of import into tomato fruits. Ber. Dtsch. Bot. Ges., 93, 315-325.
Ho L.C., 1989. Environmental effects on the diurnal accumulation of 45 Ca by young fruit and leaves of tomato plants. Ann.Bot., 63, 282-288.
Ho L.C., 1995. Responses of Ca-efficient and Ca-inefficient tomato cultivars to salinity in plant growth, calcium accumulation and blossom-end rot. J. Hortic. Sci., 70, 909-918.
Ho L.C., Grange R.I. & Picken A.J., 1987. An analysis of the accumulation of water and dry matter in tomato fruit. Plant Cell Environ., 10, 157-162.
Ho L.C. et al., 1993. Uptake and transport of calcium and the possible causes of blossom-end rot in tomato. J. Exp. Bot., 44, 509-518.
Ho L.C. & White P.J., 2005. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Ann. Bot., 95(4), 571-581.
Huygens H. & Vandecasteele P., 1992. On-line CO2 analysis for determination of cold tolerance in green house plants using a temperature controlled leaf chamber. Acta Hort., 304, 143-150.
Manishi K., Fukurnoto Y. & Yoshida T., 1996. Effects of application of compost on growth and quality of tomatoes under water stress due to root-zone restriction. J. Jpn. Soc. Hortic. Sci., 67, 257-264.
Marcelis L.F.M. & Ho L.C., 1999. Blossom-end rot in relation to growth and calcium content in fruit of sweet pepper (Capsicum annuum L.). J. Exp. Bot., 50(332), 357-363.
Martinez P.F. & Roca D., 2001. Regulation of air humidity and effects on mineral levels and blossom-end rot incidence in peppers fruits. Acta Hortic., 559, 407-412.
Mix G.P. & Marschner H., 1976. Einfluss exogener und endogener Faktoren auf den Calciumgehalt von Paprika- und Bohnenfrüchten. Z. Pflanzenernaehr. Bodenkd., 5, 551-563.
Morley P.S., 1996. Calcium nutrition and the physiology of sweet pepper (Capsicum annuum L.) fruit. PhD Thesis: Leeds University (UK).
Morley P.S., Hardgrave M., Bradley M. & Pilbeam D.J., 1993. Susceptibility of sweet pepper (Capsicum annuum L.) cultivars in the calcium deficiency disorder " blossom-end rot ". ln: Fragoso M.A.C. & Van Beusichem M.L., eds. Optimization of plant nutrition. Dordrecht, The Netherlands: Kluwer Academic Publishers, 561-567.
Nonami H. et al., 1995. Blossom-end rot of tomato plants may not be directly caused by calcium deficiency. Acta Hortic., 396, 107-114.
Paiva E.A.S., Martinez H.E.P., Casali V.W.O. & Padilha L., 1998. Occurrence of blossom-end rot in tomato as a function of calcium dose in the nutrient solution and air relative humidity. J. Plant Nutr., 21, 2663-2670.
R’him T., Hamza N. & Henchi B., 2002. Effet d’une contrainte hydrique sur la croissance et le rendement chez le piment (Capsicum annuum L.). Ann. INRAT, 75, 173-190.
R’him T. & Jebari H., 2005. Effet de différents taux d’humidité de l’air sur les échanges gazeux chez quatre variétés tunisiennes de piment (Capsicum annuum L.). Ann. INRAT, 78, 45-58.
Robbins R., 1937. Relation of nutrient salt concentration to growth of the tomato and to the incidence of blossom-end rot of the fruit. Plant Physiol., 12, 21-50.
Roca D., 1996. Necrosis apical en pimiento. Memoria de actividades 94-95. Valencia, Spain: Servicio de Desarrollo Tecnologico Agrario, Conserjeria de Agricultura.
Roca D. & Martinez P.F., 1997. Pimiento. Sensibilidad varietal a la necrosis apical. Memoria de Actividades 96-97. Valencia, Spain: Servicio de Desarrollo Tecnologico Agrario, Conserjeria de Agricultura.
Sanchez Conde M.P. & Felipe M.R., 1983. Composiciôn mineral y estudio estructural de los tejidos sanos y afectados de podedumbre apical en dos variedades de tomate (Lycopersicum esculentum L.). Ann. Edafol. Agrobiol., 42, 831-846.
Saure M.C., 2001. Blossom-end rot of tomato (Lycopersicon esculentum Mill.) a calcium or a stress related disorder. Sci. Hortic., 20, 193-208.
Tromp J., 1979. The intake curve for calcium into apple fruits under various environmental conditions. Commun. Soil Sci. Plant Anal., 10, 325-335.
Wada T., Ikeda H., Ikeda & Furukawa H., 1996. Effects of foliar application of calcium solutions on the incidence of blossom-end rot of tomato fruit. J. Jpn. Soc. Hortic. Sci., 65, 553-558.
Wiersum L.K., 1966. Calcium content of fruits and storage tissues in relation to the mode of water supply. Acta Bot. Neerl., 15, 406-418.
Willumsen J., Petersen K.K. & Kaack K., 1996. Yield and blossom-end rot of tomato as affected by salinity and cation activity ratios in the root zone. J. Hortic. Sci., 71, 81-98.
Pour citer cet article
A propos de : Thouraya Rhim
Institut National de la Recherche Agronomique de Tunisie (INRAT). Laboratoire d’Horticulture. Rue Hédi Karray. TN-2049 Ariana (Tunisie). E-mail : rhim.thouraya@iresa.agrinet.tn.
A propos de : Hager Jebari
Institut National de la Recherche Agronomique de Tunisie (INRAT). Laboratoire d’Horticulture. Rue Hédi Karray. TN-2049 Ariana (Tunisie).
