


- Accueil
- Volume 28 (2024)
- Numéro 2
- Miel de jujubier d'Algérie : analyse mélissopalynologique et caractères morphologiques du pollen


Visualisation(s): 2405 (35 ULiège)
Téléchargement(s): 219 (1 ULiège)

Miel de jujubier d'Algérie : analyse mélissopalynologique et caractères morphologiques du pollen

Document(s) associé(s)
Version PDF originaleRésumé
Description du sujet. En Algérie, le miel de jujubier a une importance socioéconomique et thérapeutique. En l’absence de normes de qualité, il est souvent exposé aux pratiques frauduleuses sur son appellation d’origine et sa qualité.
Objectifs. Identifier les caractéristiques polliniques, déterminer le spectre pollinique permettant de spécifier les miels de jujubier d’Algérie ainsi que les caractères morphologiques du pollen de Ziziphus lotus (L.) Lam.
Méthode. Quarante échantillons de miel de Z. lotus ont été soumis aux analyses polliniques qualitative et quantitative selon les méthodes préconisées par la commission internationale de botanique apicole. La description morphologique du pollen a été réalisée par microscopie électronique à balayage.
Résultats. Les miels de jujubier appartiennent aux classes II et III de Maurizio, ils ne renferment aucun indicateur de miellat. Cinquante-quatre taxons de pollen appartenant à 35 familles ont été identifiés. Le nombre de types polliniques par échantillon varie de 16 à 37. Tous les miels analysés ont une dominance de pollen de jujubier supérieure à 46 %, avec un pourcentage moyen de 65,2 % ± 13,3. Le pollen de Peganum harmala L. est omniprésent dans les miels. La combinaison en pollen de P. harmala, Retam raetam, des genres Euphorbia et Brassica forme, ainsi que certaines espèces d’astéracées des genres Centaurea, Echinops, Calendula et Carduus type, pourrait caractériser les miels de jujubier d'Algérie. Le pollen de Z. lotus est tricolporé avec de longs sillons méridiens, isopolaire à symétrie radiale, triangulaire en vue polaire, majoritairement de taille médium et de forme prolate. L’ornementation de l’exine est de type rugulé.
Conclusions. Cette étude permet de construire une base de données pollinique référentielle des miels de jujubier d’Algérie.
Abstract
Algerian jujube honey: melissopalynological analysis and morphological characteristics of pollen
Description of the subject. In Algeria, jujube honey has a socio-economic and therapeutic importance. In the absence of quality standards, it is often exposed to fraudulent practices in its designation of origin and quality.
Objectives. To identify the characteristics of pollen and to determine the pollen spectrum, making it possible to specify Algerian jujube honeys, as well as the morphological characteristics of the pollen of Ziziphus lotus (L.) Lam.
Method. Forty samples of Z. lotus honey were subjected to qualitative and quantitative pollen analyses according to the methods recommended by the International Commission of Bee Botany. The morphological description of Z. lotus pollen was performed by scanning electron microscopy.
Results. The jujube honeys belong to Maurizio’s class II and III; they do not contain any honeydew indicator. Fifty-four pollen taxa belonging to 35 families were identified. The number of pollen types per sample varied from 16 to 37. All the honey samples analyzed had a jujube pollen dominance of over 46%, with an average percentage of 65.2% ± 13.3. The pollen of Peganum harmala L. was found to be ubiquitous in the honey samples. The combination of pollen from P. harmala, Retama raetam, the Euphorbia and Brassica form genera, as well as some Asteraceae species from the genera of Centaurea, Echinops, Calendula and Carduus type, were found to characterize the Algerian jujube honey samples studied. The pollen of Z. lotus is tricolporate with long meridian colpi, isopolar with radial symmetry, triangular in polar view, mostly medium-sized and prolate-shaped. The exine ornamentation is rugulated.
Conclusions. This study enables the construction of a referential pollen database of jujube honeys from Algeria.
Table des matières
Reçu le 19 juillet 2023, accepté le 4 avril 2024, mis en ligne le 13 mai 2024.
Cet article est distribué suivant les termes et les conditions de la licence CC-BY (http://creativecommons.org/licenses/by/4.0/deed.fr)
1. Introduction
1L’Algérie, de par sa situation géographique, la complexité de ses reliefs, son hétérogénéité climatique et édaphique, est considérée comme extrêmement riche en patrimoine floristique (Véla & Benhouhou, 2007). Cette biodiversité florale abrite des ressources naturelles importantes pour l’alimentation de l’abeille tellienne, Apis mellifera intermissa, permettant ainsi la production de plusieurs miels monofloraux et polyfloraux. Les miels monofloraux sont issus principalement d’une miellée d’une seule espèce végétale donnée, ils sont dotés de propriétés physico-chimiques et organoleptiques caractéristiques et ont ainsi une valeur commerciale plus importante que les miels polyfloraux (Persano Oddo & Piro, 2004). Le miel de jujubier, autrement appelé « Sedra », fait partie des miels monofloraux les plus chers au monde (Zerrouk et al., 2018). Il occupe une place notable, très appréciée sur le plan thérapeutique et gustatif. Il est connu par sa richesse en phénols et flavonoïdes, ses propriétés antioxydantes (Khallouki et al., 2020), antimicrobiennes (Fahim et al., 2014) et son effet protecteur contre les troubles hépatiques liés à l'alcoolisme chronique (Cheng et al., 2014). Ce type de miel est produit dans plusieurs pays du monde tels que le Yémen (Alzubier & Okechukwu, 2011), l’Algérie (Zerrouk et al., 2018), la Chine (Cheng et al., 2019), l'Iran (Faal et al., 2019) et le Maroc (Khallouki et al., 2020). Le jujubier est un arbre ou arbuste principalement réparti dans les régions subtropicales et tropicales. Le genre Ziziphus Mill. renferme environ 170 espèces (Islam & Simmons, 2006), dont cinq sont présentes en Afrique du Nord, à savoir Z. lotus (L.) Lam., Z. mauritiana Lam., Z. mucronata Willd., Z. spina-christi (L.) Desf. et Z. jujuba Mill. Ces deux dernières espèces sont indiquées, respectivement, naturalisées et cultivées en Algérie. Ziziphus lotus est l’unique espèce spontanée (Dobignard & Chatelain, 2013). Selon la classification d'Emberger (1955), Z. lotus est distribué, principalement, dans les bioclimats méditerranéens semi-arides et arides (Nasri-Ayachi & Nabli, 2009). En Algérie, l’espèce est répartie dans les secteurs du Sahara septentrional, central et méridional (Quézel & Santa, 1962). La floraison du jujubier suscite un intérêt particulier des apiculteurs algériens, ils transhument massivement leurs colonies d’abeilles pour produire le miel de jujubier. Vu son importance socio-économique, ce type de miel est confronté à plusieurs pratiques frauduleuses sur son appellation d’origine et sa qualité. Ces problèmes sont courants et rencontrés dans d’autres pays (Tsagkaris et al., 2021; Bouddine et al., 2022), surtout en l’absence de cadre normatif de référence et de législation. Les analyses polliniques qualitatives et quantitatives contribuent étroitement à la connaissance des interrelations entre l’abeille et la plante butinée, et fournissent des données scientifiques probantes permettant l’identification et la caractérisation des miels (Louveaux & Abed, 1984). Elles constituent alors un outil indispensable pour toute démarche visant à préserver la qualité des miels ainsi que leurs origines géographique et botanique. L'analyse pollinique a été la première méthode utilisée pour déterminer l'origine d'un miel. La détermination de l’origine géographique est basée sur l’ensemble des espèces du spectre pollinique (Battesti & Goeury,1992). La détermination de l’origine botanique, quant à elle, repose sur les fréquences relatives des types de pollen des espèces nectarifères. En général, un miel est dit monofloral si la fréquence relative du pollen est supérieure à 45 %. Cependant, en raison des nombreux types de pollen sur- ou sous-représentés, les pourcentages de pollen peuvent varier considérablement entre différents miels monofloraux (Louveaux et al., 1978 ; Von Der Ohe et al., 2004). Les études menées sur les miels du jujubier ont été basées sur les propriétés physico-chimiques, antioxydantes et antibactériennes (Haderbache et al., 2013; Cheng et al., 2019; Khallouki et al., 2020; Zhang et al., 2020). Cependant, il existe peu de travaux sur leur caractérisation mélissopalynologique. En l’occurrence, des investigations ont été faites par Hegazi et al. (2022) sur 794 échantillons commercialisés sur le marché saoudien et importés de 12 pays. D’autres recherches portant sur deux régions algériennes steppiques, à savoir Laghouat et Djelfa, ont été publiées par Zerrouk et al. (2018, 2021) et Mekious et al. (2015, 2020). En mélissopalynologie, la microscopie électronique à balayage (MEB) s’avère également utile en taxonomie pour l'identification et la comparaison morphologique des grains de pollen. La résolution de la MEB offre une vision claire de la texture fine de la surface du pollen, permettant ainsi une distinction entre les différents taxons (Saklani & Mattu, 2020). Pour autant que nous le sachions, hormis le travail réalisé en Tunisie et publié par Nasri-Ayachi & Nabli (1995), aucune étude antérieure n’a été conduite sur l’identification des caractères morphologiques du pollen de Z. lotus d’Algérie.
2Les objectifs de notre travail visent l’identification du spectre pollinique des miels de jujubier d’Algérie et la caractérisation morphologique du pollen de Ziziphus lotus (L.) Lam.
2. Matériel et méthodes
2.1. Échantillons de miel
3Notre étude a été réalisée sur 40 échantillons de miel de jujubier produits dans 10 wilayas localisées en zones bioclimatiques diversifiées semi-arides, arides et sahariennes (Figure 1). Les échantillons ont été collectés auprès de 32 apiculteurs durant deux années de production, 2020 et 2021. Ils ont été conservés dans des pots en verre à fermeture hermétique et à une température inférieure à 4 °C.
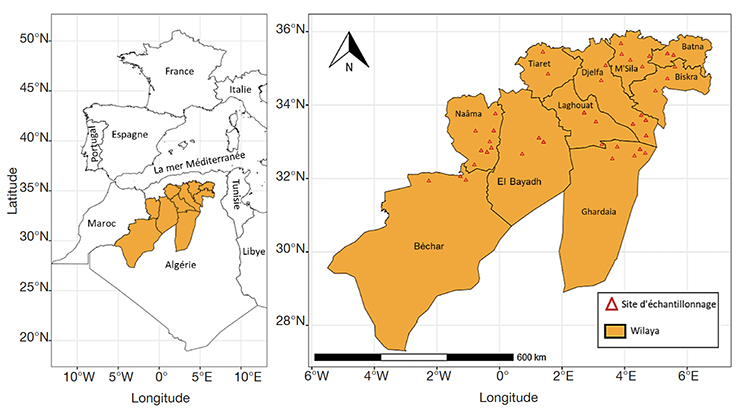
Figure 1. Zones de collecte des échantillons de miel — Honey samples collection areas.
2.2. Analyse pollinique quantitative
4L’analyse pollinique quantitative consiste à déterminer la richesse pollinique des miels. Il s’agit de calculer le nombre absolu d’éléments microscopiques par unité de poids de miel, en comptant le nombre de grains de pollen (GP) et les indicateurs de miellat contenus dans 10 g de miel au moyen d’un microscope optique et selon la méthode Maurizio (1939) citée par Louveaux et al. (1978). Le principe est d’extraire par une première centrifugation pendant 10 min à 2 500 tours·min-1, le sédiment d’une solution de miel (10 g de miel dans 20 ml d'eau distillée chaude dont la température ne dépasse pas 40 °C). De l'eau distillée est ajoutée au culot puis centrifugée à nouveau pendant 5 min à 2 500 tours·min-1 pour dissoudre entièrement les cristaux de sucre. Après décantation et élimination du liquide surnageant, le culot est monté sur une lame dans la glycérine gélatinée puis examiné au microscope optique.
5Le miel est répertorié dans l'une des cinq classes de représentativité pollinique de Maurizio (Louveaux et al., 1978 ; Von Der Ohe et al., 2004). La classe I comprend les miels monofloraux au pollen sous- représenté, pauvres en pollen (GP/10 g < 20 × 103). La classe II inclut la plupart des miels de fleurs et ceux mélangés de miel de miellat, avec une représentativité pollinique normale (GP/10 g de 20 × 10 à 100 × 103). Les miels de la classe III regroupent les miels de miellat et les miels monofloraux avec un pollen surreprésenté (GP/10 g de 100 × 103 à 500 × 103), la classe IV comprend les miels monofloraux à forte surreprésentation pollinique et certains miels pressés (GP/10 g de 500 × 103 à 106) et la classe V comprend les miels pressés (GP/10 g > 106 pollen).
2.3. Analyse pollinique qualitative
6L’analyse pollinique qualitative consiste à identifier et à dénombrer les différents types de pollen contenus dans les miels. Ainsi, les fréquences polliniques exprimées en pourcentage relatif ont été calculées par rapport au nombre total de grains de pollens comptés. Le dénombrement et l’identification des types polliniques ont été réalisés en suivant la méthode décrite par Louveaux et al. (1978) et Von Der Ohe et al. (2004). Ainsi, 10 g de miel ont été dissous dans 10 ml d'eau distillée acidulée à 5 % d’acide sulfurique puis centrifugés en deux temps à 2 500 tours·min-1 pour pouvoir dissoudre les cristaux de sucres restants. Le sédiment est récupéré et monté sur une lame dans la glycérine gélatinée pour l’observation sous microscope photonique. Les classes de fréquences relatives ont été déterminées sur un nombre total égal à 800 grains de pollen comptés. Quatre classes de fréquences polliniques ont été différenciées selon la nomenclature mélissopalynologique établie par la commission internationale de botanique apicole, l’U.I.S.B (Union Internationale des Sciences Biologiques), telle qu’explicitée par Louveaux et al. (1978) :
7- pollens prédominants ou dominants (˃ 45 %) ;
8- pollens secondaires ou d’accompagnement (16 à 45 %) ;
9- pollens tertiaires ou isolés importants (3 à 15 %) ;
10- pollens rares ou isolés (˂ 3 %).
11L'identification des types de pollen est réalisée par la comparaison des caractères morphologiques et des dimensions des grains de pollen observés au microscope photonique dans nos échantillons avec ceux des microphotographies des pollens de référence établies par Ricciardelli D’Albore (1998). Nous avons aussi utilisé quelques lames et micrographies de références que nous avons effectuées durant nos études antérieures. Il est important de souligner que l'identification précise de l’espèce de certains grains de pollen d'Apiaceae, de Brassicaceae, de Casuarinaceae et de Fabaceae n'était pas possible ; ils ont été mentionnés selon la forme du genre (ex : Sinapis form, Casuarina form). Certains types polliniques ont été indiqués comme Apiaceae (other) ou Fabaceae (other) pour les distinguer des types identifiés de la même famille botanique.
12La fréquence de distribution des types polliniques est calculée en fonction du nombre de miels analysés dans lesquels ils sont retrouvés, comme indiqué par Feller-Demalsy & Parent (1989), ils sont classés en :
13- taxon très fréquent (> 50 %) ;
14- taxon fréquent (20 à 50 %) ;
15- taxon peu fréquent (10 à 20 %) ;
16- taxon rare (< 10 %).
2.4. Caractéristiques microscopiques des grains de pollen de Ziziphus lotus
17Des fleurs de Z. lotus ont été collectées dans quatre régions appartenant à des bioclimats différents, il s’agit de l’aride inférieur à hiver très froid pour la localité de Djniene Bourezg (wilaya de Naama), l’aride inférieur à hiver froid pour la localité de Loutaya (wilaya de Biskra), le saharien supérieur à hiver froid pour la localité de Beni Ounif (wilaya de Bechar) et le saharien inférieur à hiver froid pour la localité de Berriane (wilaya de Ghardaïa). Les anthères ont été isolées et séchées puis conservées dans des tubes secs à hémolyse. Extraits des anthères, les grains de pollen n’ont pas été acétolysés ; ils ont été préparés selon la technique de métallisation réalisée au laboratoire du centre de recherche et d’analyses physico-chimique (CRAPC) de Ouargla, pour mener à une description morphologique qualitative et quantitative par microscopie électronique à balayage (MEB). La surface de la préparation a été rendue conductrice en appliquant un film d’or à l'aide d'un métalliseur. L’introduction des échantillons dans une chambre MEB (ZEISS CVO 15) exposée au vide permet d'obtenir une imagerie tridimensionnelle de haute résolution. Les caractéristiques morphologiques retenues sont : la polarité, la symétrie, la vue polaire, la forme, la taille, l'aspect de l'exine, le nombre d’apertures, le diamètre de l'axe polaire (P) et de l’axe équatorial (E) et enfin, le ratio P/E. L’étude est basée sur des données obtenues à partir de 169 grains de pollen. Pour l'identification des caractéristiques morphologiques, la terminologie descriptive définie par Erdtman (1952, 1945) et Punt et al. (2007) a été suivie. La taille du pollen dépend de l’axe le plus long (P), permettant ainsi d’obtenir différentes classes de taille : très petite (˂ 10 µm), petite (10-25 µm), moyenne (25-50 µm), gros (50-100 µm), très gros (100-200 µm), gigantesque (˃ 200 µm). La forme est liée au ratio P/E, ce qui permet de définir différentes classes de forme : peroblate (˂ 0,5), oblate (0,5-0,75), sub-oblate (0,75-0,88), oblate-sphéroïdale (0,88-1), prolate-sphéroïdale (1-1,14), sub-prolate (1,14-1,33), prolate (1,33-2), perprolate (˃ 2). Les résultats ont été comparés à ceux obtenus dans d’autres régions du monde.
2.5. Analyses statistiques
18Louveaux et al. (1970) et Von Der Ohe et al. (2004) recommandent l’élimination du pollen d’espèces anémophiles et/ou dépourvues de nectaires lors du calcul des fréquences polliniques des plantes nectarifères. De ce fait, l’effet de l'élimination de ces pollens sur le pourcentage pollinique de Z. lotus a été analysé par un modèle linéaire général mixte (GLMM) avec un lien logit et une distribution d’erreur quasi-binomiale, choisi pour remédier à la sur-dispersion des données. En outre, en raison de la nature appariée des échantillons, où le pourcentage de pollen de Z. lotus a été calculé à la fois en tenant compte de tous les types polliniques présents dans le miel et après avoir retranché le pollen des plantes anémophiles et/ou dépourvues de nectaires du nombre total de pollen compté, un modèle mixte a été construit. La variable Groupe, composée de deux modalités, présence et absence du pollen d’espèces anémophiles et/ou dépourvues de nectaires, est incluse en tant qu’effet fixe, tandis que les échantillons sont considérés comme une variable à effet aléatoire, ce qui permet au modèle de prendre en compte la dépendance entre les échantillons.
19Afin d’évaluer la variation de la fréquence pollinique de Z. lotus entre les miels de jujubier de la classe II et III de Maurizio, un modèle linéaire généralisé (GLM) avec un lien logit et une distribution d’erreur quasi-binomiale a été appliqué. Le nombre des types polliniques contenu dans les miels de jujubier des classes II et III a été comparé au moyen du test t de Student non apparié. Les analyses statistiques ont été effectuées à l'aide du logiciel R 4.2.3. Le GLM a été réalisé avec la fonction glm(), qui fait partie du package de base de R, tandis que le GLMM a été réalisé avec la fonction glmer() implémentée dans le package lme4.
20Les résultats morphométriques du grain de pollen de Z. lotus, incluant les variables du diamètre de l’axe polaire, le diamètre de l’axe équatorial et le ratio P/E, ont été individuellement soumis à une analyse de la variance, ANOVA à un facteur, afin d'évaluer l'existence de différences entre les moyennes de ces variables des quatre régions à bioclimats différents. Un test post-hoc de Duncan a été appliqué pour comparer les moyennes deux à deux pour identifier des paires de régions possédant des différences à l'aide du logiciel Gen Stat Discovery Edition 3. Les résultats ont été considérés comme significatifs pour une p-valeur inférieure à 0,05.
3. Résultats
3.1. Analyses polliniques quantitative et qualitative
21L’analyse pollinique quantitative révèle une variation du nombre absolu de grains de pollen par 10 g de miel, qui varie entre 48 500 et 177 500, avec une moyenne de 104 912,82 ± 39 490,67. Nous notons que 19 échantillons (47,5 %) appartiennent à la classe II de Maurizio, indiquant une représentativité pollinique normale, avec une richesse pollinique allant de 48 500 à 96 000 GP·10 g-1. Par ailleurs, 21 échantillons (52,52 %) appartiennent à la classe III, correspondant au pollen surreprésenté, avec une richesse pollinique qui varie de 110 500 et 177 500 GP·10 g-1 (Figure 2). Nous notons également l’absence d’indicateur de miellat dans les échantillons de miels étudiés.
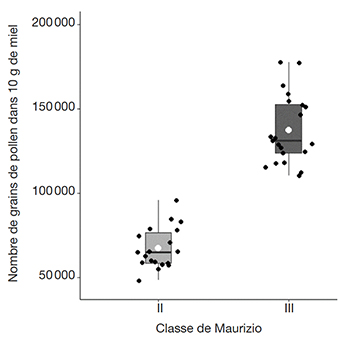
Figure 2. Analyse pollinique quantitative des échantillons de miel — Quantitative pollen analysis of honey samples.
22L’analyse pollinique qualitative (Tableau 1) met en évidence une diversité florale d'espèces spontanées et cultivées, butinées par l'abeille tellienne (A. mellifera intermissa) dans les sites d’échantillonnage de miels.
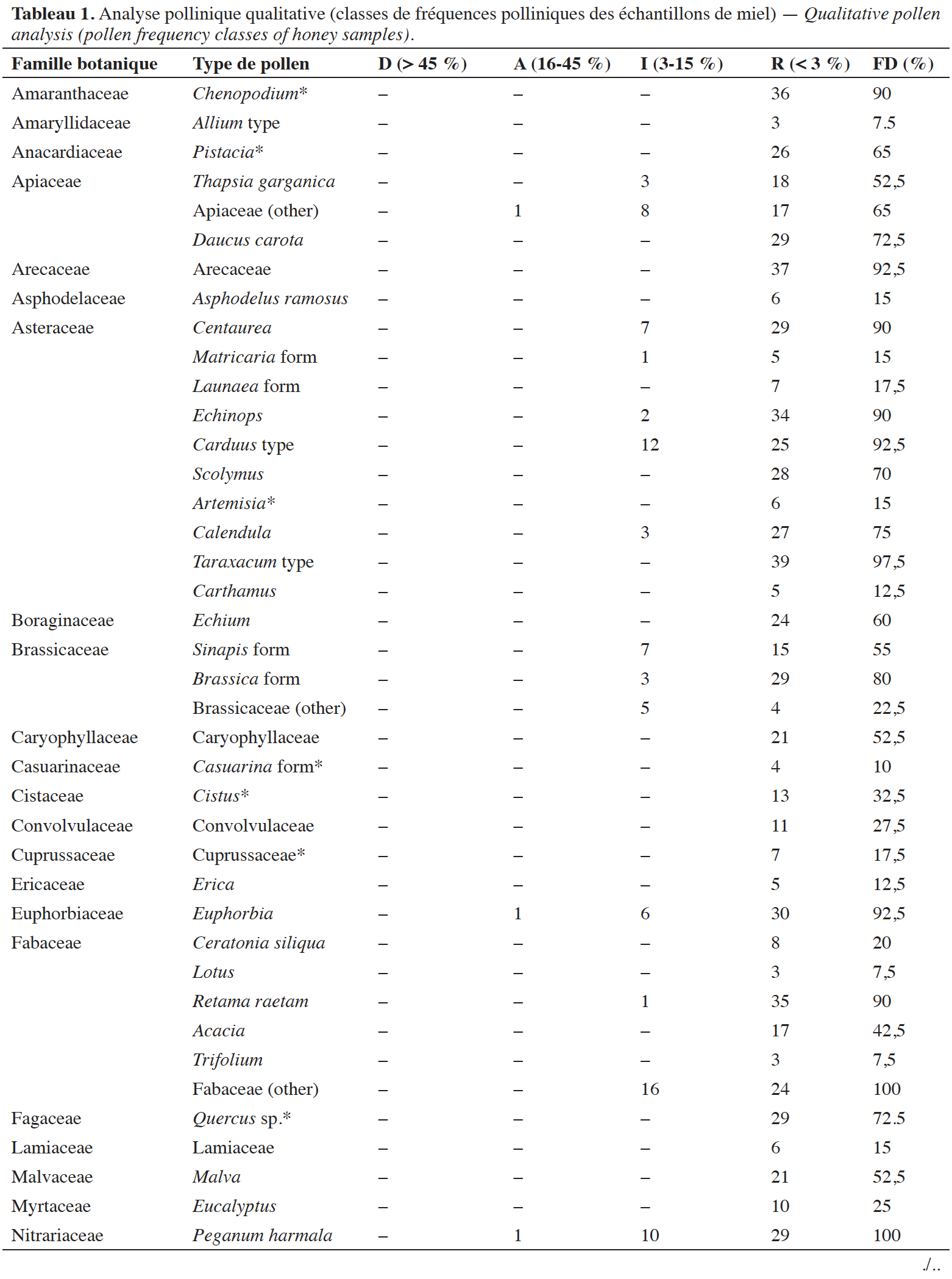

23Le nombre de types polliniques varie de 16 à 37 par échantillon de miel. Au total, 54 types polliniques ont été identifiés appartenant à 35 familles botaniques, dont les plus représentées sont les Astéracées et les Fabacées. En se référant aux travaux de Louveaux et al. (1970), Louveaux & Abed (1984), Ricciardelli D’Albore (1998) et Zerrouk et al. (2021), nous avons également relevé 12 types polliniques indiqués comme anémophiles et/ou dépourvus de nectaires. Il s’agit d’Olea europaea, Papaver rhoeas, Artemesia, Casuarina, Chenopodium, Cistus, Pistacia, Plantago, Quercus sp., Cuprussaceae, Pinaceae et Poaceae. Ils appartiennent tous à la classe des pollens rares. Concernant la fréquence de distribution des types polliniques dans les échantillons de miel analysés, nous notons que 28 types sont très fréquents, neuf sont fréquents, 12 sont peu fréquents et cinq types sont rares.
24Les pollens de R. raetam, Euphorbia, Chenopodium, Echinops, Centaurea, Carduus type, Taraxacum type, Brassica forme, Arecaceae, Poaceae ont une fréquence de distribution ≥ 80 %. Les grains de pollen de Z. lotus, P. harmala, Fabaceae (other) sont présents dans l’ensemble des miels analysés, ceux de Z. lotus sont dominants. Les types polliniques de P. harmala, d’Euphorbia et d’Apiaceae (other) sont secondaires dans trois échantillons. Le reste des types polliniques identifiés sont classés comme pollen isolé important et/ou en pollen rare.
25La fréquence pollinique relative de Z. lotus se situe entre 46,1 % et 92 %, avec une moyenne de 65,2 % ±13,3. Après élimination des types non nectarifères, la fréquence varie de 48,4 % à 93,2 % avec une teneur moyenne de 68,2 % ± 12,9.
26Les résultats du GLMM indiquent qu’après élimination du pollen des espèces anémophiles et/ou dépourvues de nectaires du total du pollen compté, la fréquence pollinique relative de Z. lotus ne démontre pas de variation statistiquement significative (p-valeur = 0,752, valeur z = -0,32) (Tableau 2).

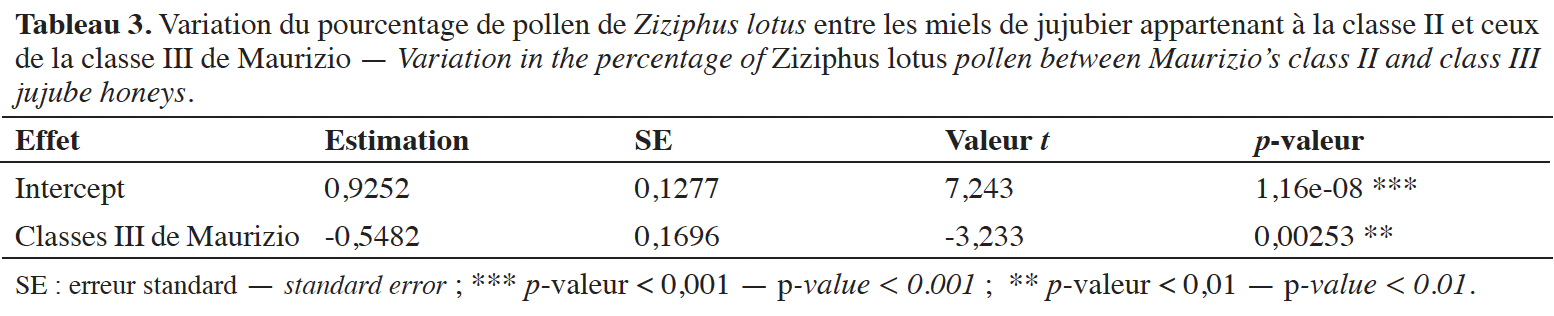
27Les résultats du GLM révèlent une variation hautement significative de la fréquence pollinique relative du Z. lotus entre les miels de jujubier de la classe II et ceux de la classe III (p-valeur = 0,00253**, ddl = 38, valeur t = -3,233). Les échantillons de miel de classe III ont une moyenne plus faible de pollen de Z. lotus (Tableau 3).
28Le test t de Student non apparié montre que le nombre de types polliniques diffère significativement (p-valeur = 0,0417, ddl = 38, valeur t = -2,1071) entre les deux groupes de miel. Les échantillons de miel de la classe II ont un nombre moyen de types de pollen inférieur à celui des miels de la classe III (Figure 3).
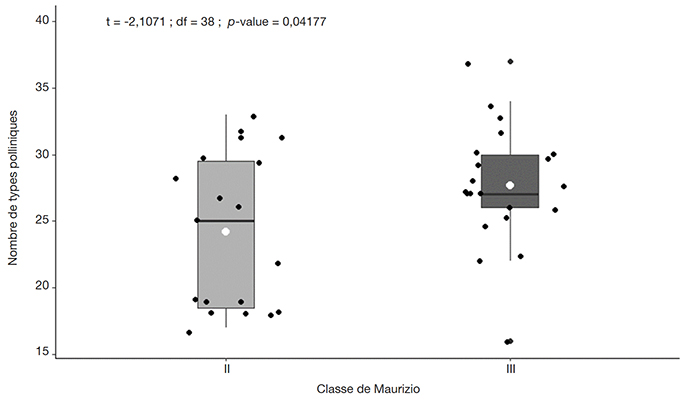
Figure 3. Nombre de types polliniques dans les échantillons de miel selon les classes II et III de Maurizio — Number of pollen types in honey samples according to Maurizio’s classes II and III.
3.2. Caractéristiques microscopiques des grains de pollen de Ziziphus lotus
29La détermination des caractères morphologiques du pollen de Z. lotus est basée sur des critères qualitatifs et quantitatifs (Figure 4 et Tableau 4). Ziziphus lotus a des grains de pollen tricolporés avec de longs sillons méridiens (longicolporés). Ils sont isopolaires à symétrie radiale, la vue polaire est triangulaire. L’ornementation de l’exine est de type rugulé.
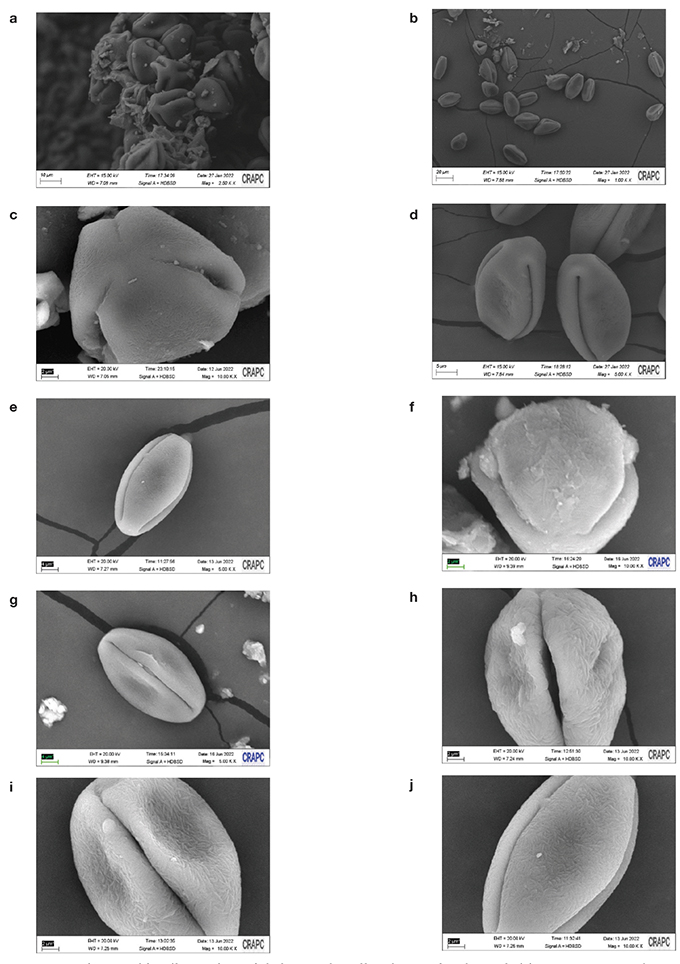
Figure 4. Micrographies électroniques à balayage de pollen de Ziziphus lotus algérien — Scanning electronmicrographs of Algerian Ziziphus lotus pollen.
a-b : forme du pollen — pollen shape, c : polaire triangulaire — triangular polar, d : vue polaire — polar view, e-f-g : vue équatoriale — equatorial view, h-i-j : ornementation d’exines — exine ornamentation.
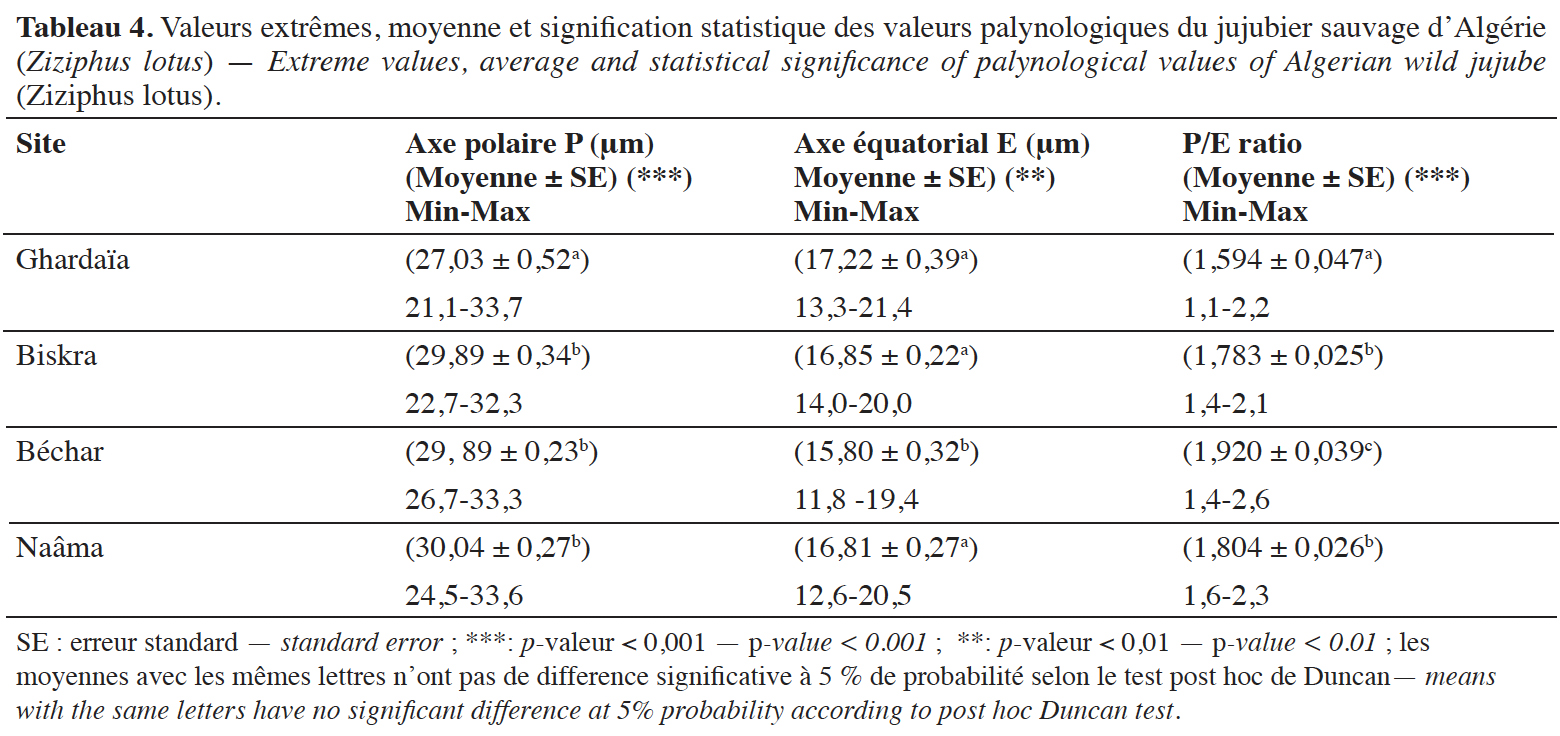
30La taille du pollen varie de 21,1 à 33,7 µm. La moyenne la plus élevée provient de la localité de Naama (30,04 µm) et la plus faible est celle de la localité de Ghardaïa (27,03 µm). Selon la classification adoptée par Erdtman (1952), la majorité des grains de pollen étudiés (92,9 %) appartiennent à la classe de pollen de taille moyenne, tandis que le reste des pollens (7,1 %) est de taille petite. Le diamètre équatorial (E) varie de 11,8 à 21,4 µm, la moyenne la plus élevée concerne les grains de pollen de la localité de Ghardaïa (17,22 µm) ; la moyenne la plus faible est celle du pollen de Béchar (15,80 µm).
31La forme du pollen fait référence au ratio P/E, qui varie de 1,1 à 2,6 et dont la plus grande et la plus faible moyenne sont enregistrées respectivement pour le pollen des localités de Béchar (1,92) et de Ghardaïa (1,59). D’après la classification d’Erdtman (1952), trois formes polliniques ont été distinguées. La forme prolate est dominante (79,3 %), suivie de la forme per-prolate (17,7 %) ; la forme sub-prolate (3 %) est rarement observée et concerne uniquement quelques grains de pollen de la localité de Ghardaïa.
32Les résultats de l'analyse de la variance ANOVA à un facteur révèlent l'existence de différences très hautement significatives entre les quatre régions à bioclimats différents pour les moyennes des deux variables : le diamètre de l'axe polaire P (p-valeur ˂ 0,001, ddl = 3, valeur F = 15,424) et le rapport P/E (p-valeur ˂ 0,001, ddl = 3, valeur F = 13,082). De plus, des différences hautement significatives sont également observées pour les moyennes de la variable du diamètre équatorial E (p-valeur ˂ 0,01, ddl = 3, valeur F = 3,937).
33Le test de comparaison multiple des moyennes indique que le diamètre de l'axe polaire des grains de pollen collectés à Biskra, Béchar et Naâma constitue un groupe homogène qui diffère de celui de Ghardaïa. Les moyennes du diamètre équatorial du pollen de Biskra, Naâma et Ghardaïa suggèrent une homogénéité qui diffère de celle du pollen provenant de Béchar. Concernant la moyenne du rapport P/E, trois groupes se distinguent : il s'agit du pollen de la localité de Ghardaïa, celui de Béchar et ceux des deux localités d’apparition homogène Biskra et Naâma.
4. Discussion
34Les miels de jujubier produits en Algérie sont caractérisés par une richesse importante en pollen avec un nombre absolu situé dans les classes II et III de Maurizio. Les miels appartenant à la classe II renferment un nombre de types polliniques moindre et une fréquence pollinique de Z. lotus élevée par rapport aux miels de la classe III. Puusepp & Koff (2014) rapportent que les variations de composition et de densité pollinique sont liées au potentiel pollinifère des différentes espèces végétales, au type de végétation entourant les ruches ainsi qu'aux variations des conditions climatiques.
35La diversité en types polliniques des miels analysés est justifiée par l’échelonnement de la floraison du jujubier (de mai à juin) qui coïncide avec celle de nombreuses espèces spontanées et cultivées, et par l’élargissement du rayon de butinage des abeilles pour l’approvisionnement en nectar et en pollen. La présence de pollen d’espèces cultivées dans le spectre pollinique (Olea europaea, Citrus, Prunus et Arecaceae) reflète le développement de l'arboriculture dans les espaces steppiques et sahariens, suite au plan national de développement agricole (PNDA) mis en œuvre par les pouvoirs publics depuis 1998 (Khaldi, 2014). Le pollen de P. harmala a été cité comme dominant parmi les miels du Constantinois algérien (Louveaux & Abed, 1984) et ceux de la Tunisie (Ben Haj Jilani et al., 2008). L'omniprésence du pollen de P. harmala a été mentionnée par Bahloul et al. (2022) dans les miels de Laghouat (Secteur de l’Atlas saharien et secteur du Sahara septentrional) ; il figure en dominance et/ou en accompagnement dans le spectre pollinique. Peganum harmala est une plante commune des hauts plateaux, du Sahara septentrional et des montagnes du Sahara central, elle se développe dans les sols sableux et peu nitratés (Ozenda, 2004). Dans les miels Bayadh (Secteur des hauts plateaux et Secteur de l’Atlas saharien), le pollen d’Euphorbia atteint la classe du pollen dominant ou d’accompagnement (Haderbache et al., 2013). Le genre Euphorbia comprend 150 espèces, il est important au Sahara septentrional et en Europe (Ozenda, 2004). Louveaux & Abed (1984) soulignent l'importance des Apiaceae dans les miels d'Afrique du Nord. La représentation importante des familles des Fabaceae et des Asteraceae a été signalée aussi dans des études menées en Algérie (Hamel et al., 2019 ; Ghorab et al., 2021) et dans la région centrale du Rif du Maroc d'Azilal et de Beni Mellal (Terrab et al., 2022).
36Comme dans la plupart des miels monofloraux, le pollen de jujubier se trouve dominant avec une fréquence de plus de 46 %. Des résultats similaires en Algérie ont été obtenus par Zerrouk et al. (2021) où la dominance du pollen de Z. lotus varie de 45,3 à 93,4 % dans les miels de Laghouat et Djelfa. Mekious et al. (2015) ont rapporté, dans leur étude sur des miels de jujubier de Djelfa, un pourcentage pollinique qui varie entre 45,75 et 97,12 %.
37La combinaison des types polliniques P. harmala, R. raetam, Euphorbia, Centaurea, Echinops, Calendula et Carduus type, et Brassica forme pourrait spécifier les miels de jujubier d'Algérie. Ces types polliniques ont à la fois une distribution de présence élevée et des fréquences relatives importantes, permettant de les classer en pollens secondaires ou tertiaires. Zerrouk et al. (2021) ont trouvé l’association des types polliniques de P. harmala, Thapsia garganica, Limonium bonduellei, Muscari comosum, Launaea, Carthamus et plusieurs Asteraceae tels que des espèces des genres Centaurea, Taraxacum, Carduus, Artemisia et Matricaria, comme combinaison pollinique caractéristique des miels de jujubier de Djelfa et de Laghouat. Pour ces mêmes régions, Mekious et al. (2020) ont suggéré la combinaison pollinique composée d’Euphorbia bupleuroides, P. harmala, R. raetam, T. garganica, Asteraceae, Brassicaceae et Poaceae. Les grains de pollen qui enregistrent une présence constante sont ceux de P. harmala et des Asteraceae. Le spectre pollinique du miel de jujubier d’Algérie est différent de celui obtenu par Hegazi et al. (2022) pour le miel de Z. lotus d’Égypte, qui est composé de pollen de Brassica tournefortii Gouan, d’Oryza meyeriana et de Zea mays.
38Le pollen de Z. lotus présente des caractères morphologiques uniformes au niveau de la symétrie, la polarité, l’aperture, la vue polaire et l’ornementation de l’exine. La variabilité a été enregistrée au niveau de la dimension de l'axe polaire P, du diamètre équatorial et du rapport P/E (Tableau 3). Ces caractères morphologiques sont généralement similaires à ceux du pollen de la famille des Rhamnaceae en termes d’apertures (tricolporées) et de taille qui varie entre les tailles petite et moyenne. Cependant, la forme et les types d'ornementation montrent une nette hétérogénéité. Le pollen de R. alaternus du bassin méditerranéen se caractérise par une forme sub-oblate et une ornementation fovéolée (Ricciardelli D’Albore, 1998). Le pollen de Z. jujuba Mill. de la région de Yang en Myanmar est de forme sub-oblate et d’ornementation légèrement réticulée (Myint, 2019), tandis que celui de l'Inde est de forme prolate-sphéroïdale et d'ornementation rugulée (Dinesh, 2018). Le pollen du cultivar iranien de Z. jujuba Mill. est de forme oblate-sphéroïdale et d'ornementation irrégulièrement striée (Rouhakhsh et al., 2014). Quant au pollen de Z. lotus, nos résultats montrent une forme majoritairement prolate et une ornementation rugulée, ce qui diffère des résultats obtenus par Nasri-Ayachi & Nabli (1995) en Tunisie, où la forme oblate et une ornementation de l’exine striato-rugulée ont été observées.
39Les variations biométriques des grains de pollen ont été expliquées par plusieurs auteurs. Flenley (2003) souligne que les traitements chimiques au laboratoire, tels que l’acétolyse et les milieux de montage, modifient considérablement la taille du pollen. Fatmi et al. (2020) rapportent que les facteurs biotiques et abiotiques tels que la température, l'altitude, les pollinisateurs peuvent provoquer ces variations. Belhadj et al. (2007) ainsi que Khouatmiani et al. (2021) ont montré que la taille peut varier, également, en plus de l’altitude et de la température, en fonction de plusieurs autres facteurs géoclimatiques tels que l’aridité, la latitude, la longitude et la pluviométrie du site, respectivement, chez le pollen de Pistacia atlantica et d’Olea europaea. En effet, des échantillons prélevés de plusieurs stations variant de l’aride à l’humide exhibaient des formes et des tailles variées des grains de pollen des deux espèces étudiées (P. atlantica et O. europaea). Dans ces contrées arides et semi-arides, le jujubier joue le rôle de plante nurse, protectrice pour P. atlantica. Il est intéressant de signaler que le jujubier (Z. lotus) et le pistachier de l’Atlas (P. atlantica) partagent les mêmes bioclimats et donc l’on peut supposer qu’ils répondent de la même manière aux contraintes environnementales.
5. Conclusions
40L’étude mélissopalynologique menée sur les miels de jujubier d’Algérie a permis de révéler un spectre riche en types polliniques. Selon la nomenclature mélissopalynologique internationale, tous les miels analysés sont monofloraux avec une fréquence pollinique de Z. lotus supérieure à 46 %. La présence de certains types dans le spectre pollinique global, comme P. harmala, R. raetam, Euphorbia, Centaurea, Echinops, Calendula et Carduus type, et Brassica forme, pourrait constituer un gage de qualité permettant d’une part, de caractériser les miels de jujubier d’Algérie et de les distinguer de ceux de même origine botanique mais produits dans d’autres pays et, d’autre part, de détecter les éventuelles fraudes sur l’origine géographique. Des études similaires doivent être effectuées sur plusieurs années afin de constituer une base de données référentielle qui permettra de déterminer, avec exactitude, les combinaisons polliniques caractéristiques du miel de jujubier d’Algérie. L’étude de l'ultra-structure du pollen de Z. lotus a mis en évidence une homogénéité des caractéristiques morphologiques en termes de symétrie, polarité, aperture, vue polaire et ornementation de l’exine. La variabilité a été enregistrée au niveau de la taille et de la forme. Par conséquent, un échantillonnage suffisant mené sur plusieurs sites peut aboutir à des descriptions morphologiques décisives des grains de pollen et une identification de la variabilité intraspécifique.
Remerciements
41Les auteurs remercient les apiculteurs professionnels locaux pour leur disponibilité et la fourniture d'échantillons de miel, le personnel de la conservation des forêts de la wilaya de Naama pour le soutien technique lors des sorties sur le terrain, ainsi que le conservateur au Jardin botanique de la Ville de Genève, Monsieur Cyril Châtelain, pour son aide dans l'identification et la systématique des espèces végétales.
Bibliographie
Alzubier A.A. & Okechukwu P.N., 2011. Investigation of anti-inflammatory, antipyretic and analgesic effect of Yemeni Sidr honey. World Acad. Sci. Eng. Technol., 5(8), 420-425.
Bahloul R., Zerrouk S. & Chaibi R., 2022. Pollen analysis of honey from Laghouat region (Algeria). Grana, 61(6), 471-480, doi.org/10.1080/00173134.2022.2126726
Battesti M.J. & Goeury C., 1992. Efficacité de l’analyse mélitopalynologique quantitative pour la certification des origines géographique et botanique des miels: le modèle des miels corses. Rev. Palaeobot. Palynol., 75(1-2), 77-102, doi.org/10.1016/0034-6667(92)90151-6
Belhadj S. et al., 2007. Pollen morphology and fertility of wild Atlas pistachio (Pistacia atlantica Desf., Anacardiaceae). Grana, 46(3), 148-156, doi.org/10.1080/00173130701520310
Ben Haj Jilani I. et al., 2008. Physicochemical properties and pollen spectra of honeys produced in Tunisia (Southwest of Kef). Apiacta, 43, 38-48.
Bouddine T. et al., 2022. Organic honey from the Middle Atlas of Morocco: physicochemical parameters, antioxidant properties, pollen spectra, and sugar profile. Foods, 11, 3362, doi.org/10.3390/foods11213362
Cheng N. et al., 2014. Antioxidant properties of jujube honey and its protective effects against chronic alcohol-induced liver damage in mice. Food Function, 5(5), 900-908, doi.org/10.1039/c3fo60623f
Cheng N. et al., 2019. Jujube honey induces apoptosis in human hepatocellular carcinoma HepG2 cell via DNA damage, p53 expression, and caspase activation. J. Food Biochem., 43(11), 1-13, doi.org/10.1111/jfbc.12998
Dinesh B., 2018. Pollen biology and morphology of Ziziphus jujuba Mill. (Rhamnaceae). Int. J. Acad. Res. Dev., 3(2), 819-821.
Dobignard A. & Chatelain C., 2013. Index synonymique de la flore d’Afrique du Nord. Vol. 5 : Dicotyledoneae : Oleaceae à Zygophyllaceae. Genève, Suisse : Ville de Genève, Éditions des Conservatoire et Jardins botaniques.
Erdtman G., 1945. Pollen morphology and plant taxonomy IV. Labiatae, Verbenaceae and Avicenniaceae. Svenk Bot. Tidskr., 39, 279-285.
Erdtman G., 1952. Pollen morphology and plant taxonomy: angiosperm. Waltham, MA, USA: Chronica Botanica Co.; Stockholm: Almqvist and Wiksell ed.
Faal S., Loghavi M. & Kamgar S., 2019. Physicochemical properties of Iranian ziziphus honey and emerging approach for predicting them using electronic nose. Measurement, 148, 106936, doi.org/10.1016/j.measurement.2019.106936
Fahim H. et al., 2014. Physico-chemical analysis and antimicrobial potential of Apis dorsata, Apis mellifera and Ziziphus jujube honey samples from Pakistan. Asian Pac. J. Trop. Biomed., 4(8), 633-641, doi.org/10.12980/APJTB.4.2014APJTB-2014-0095
Fatmi H. et al., 2020. Pollen morphological variability correlates with a large-scale gradient of aridity. Web Ecol., 20, 19-32, doi.org/10.5194/we-20-19-2020
Feller-Demalsy M.J. & Parent J., 1989. Analyse pollinique des miels de l’Ontario, Canada. Apidologie, 20(2), 127-138, doi.org/10.1051/apido:19890203
Flenley J., 2003. Some prospects for lake sediment analysis in the 21st century. Quat. Int., 105, 77-80, doi.org/10.1016/s1040-6182(02)00153-2
Ghorab A. et al., 2021. Characteristics of honey from Babors Kabylia’s Region (Algeria). Foods, 10(225), 1-17, doi.org/10.3390/foods10020225
Haderbache L., Bousdira M. & Mohammedi A., 2013. Ziziphus lotus and Euphorbia bupleuroides Algerian honeys. World Appl. Sci. J., 24(11), 1536-1543, doi.org/10.5829/idosi.wasj.2013.24.11.7525
Hamel T., Bellili M., Meddad-Hamza A. & Boulemtafes A., 2019. Nouvelle contribution à l’étude de la flore mellifère et caractérisation pollinique de miels de la Numidie (Nord-Est algérien). Livest. Res. Rural Dev., 31(11), 1-10.
Hegazi A.G. et al., 2022. Characterization of sidr (Ziziphus spp.) honey from different geographical origins. Appl. Sci., 12(18), 9295, doi.org/10.3390/app12189295
Islam M.B. & Simmons M.P., 2006. A thorny dilemma: testing alternative intrageneric classifications within Ziziphus (Rhamnaceae). Syst. Bot., 31(4), 826-842, doi.org/10.1600/036364406779695997
Khaldi A., 2014. La gestion non-durable de la steppe algérienne. VertigO, doi.org/10.4000/vertigo.15152
Khallouki F. et al., 2020. HPLC-ESI-MS and GC-EI-MS identification and quantitation of polyphenolics and alkaloids in Moroccan jujube honeys. J. Apic. Sci., 64(2), 287-299, doi.org/10.2478/jas-2020-0029
Khouatmiani K. et al., 2021. Variability of eight Algerian oleaster ecotypes (Olea europaea subsp. europaea var. sylvestris [Mill.] Lehr): pollen and exine morphology in relation to geo-climatic effect. Grana, 60(4), 256-270, doi.rg/10.1080/00173134.2020.1815832
Louveaux J. & Abed L., 1984. Les miels d’Afrique du Nord et leur spectre pollinique. Apidologie, 15(2), 145-170, doi.org/10.1051/apido:19840205
Louveaux J., Maurizio A. & Vorwohl G., 1970. Commission internationale de botanique apicole de l’U.I.S.B. : les méthodes de la mélisso-palynologie. Apidologie, 1(2), 211-227.
Louveaux J., Maurizio A. & Vorwohl G., 1978. Methods of melissopalynology. Bee World, 59(4), 139-157, doi.org/10.1080/0005772x.1978.11097714
Mekious S. et al., 2015. Caractérisation des miels produits dans la région steppique de Djelfa en Algérie. Biotechnol. Agron. Soc. Environ., 19(3), 219-229.
Mekious S., Daoud N. & Belhadj S., 2020. Caractéristiques mélissopalynologiques et contenu phénolique du miel de Ziziphus lotus d’Algérie. Rev. Agrobiologia, 10(2), 2220-2228.
Myint D.N.Z., 2019. Study on pollen morphology of entomophilous plants in Dagon University Campus. Commemoration of 25th Anniversary Silver Jubilee Research Journal, 9(2), 328-334.
Nasri-Ayachi M.B. & Nabli M.A., 1995. Pollen wall ultrastructure and ontogeny in Ziziphus lotus L. (Rhamnaceae). Rev. Palaeobot. Palynol., 85, 85-98, doi.org/10.1016/0034-6667(94)00084-W
Nasri-Ayachi M.B. & Nabli M.A., 2009. Floral biology study of Ziziphus lotus L. Acta Hortic., 840, 337-342, doi.org/10.17660/ActaHortic.2009.840.46
Ozenda P., 2004. Flore et végétation du Sahara. Paris : Éditions du CNRS.
Persano Oddo L. & Piro R., 2004. Main European unifloral honeys: descriptive sheets. Apidologie, 35(Suppl. 1), doi.org/10.1051/apido:2004049
Punt W. et al., 2007. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol., 143(1-2), 1-81, doi.org/10.1016/j.revpalbo.2006.06.008
Puusepp L. & Koff T., 2014. Pollen analysis of honey from the Baltic region, Estonia. Grana, 53(1), 54-61, doi.org/10.1080/00173134.2013.850532
Quézel P. & Santa S., 1962. Nouvelle flore de l’Algérie et des régions désertiques méridionales. Paris : Éditions du CNRS, 1170.
Ricciardelli D’Albore G., 1998. Mediterranean melissopalynology. Perugia, Italy : Università degli studi di Perugia, Facoltà di agraria, Istituto di entomologia agraria.
Rouhakhsh H., Davarynejad G., Abedi B. & Rahemi M., 2014. Pollen grain morphological characteristics of ten Iranian jujube (Ziziphus jujuba) cultivars by using scanning electron microscopy. Adv. Environ. Biol., 8(5), 1333-1338.
Saklani S. & Mattu V.K., 2020. Scanning electron microscopic study on pollens of 8 bee floral resources from Kangra Hills, Himachal Pradesh, India. Int. J. Biotech Trends Technol., 10(1), 67-71, doi.org/10.14445/22490183/ijbtt-v10i1p611
Terrab A. et al., 2022. A palynological and geographical characterization of labeled resin spurge honey: Euphorbia resinifera. Palynology, 46(4), 1-7, doi.org/10.1080/01916122.2021.1933639
Tsagkaris A.S. et al., 2021. Honey authenticity: analytical techniques, state of the art and challenges. RSC Adv., 11(19), 11273-11294, doi.org/10.1039/d1ra00069a
Véla E. & Benhouhou S., 2007. Évaluation d’un nouveau point chaud de biodiversité végétale dans le Bassin méditerranéen (Afrique du Nord). C.R. Biol., 330(8), 589-605, doi.org/10.1016/j.crvi.2007.04.006
Von Der Ohe W. et al., 2004. Harmonized methods of melissopalynology. Apidologie, 35, S18-S25, doi.org/10.1051/apido:2004050
Zerrouk S., Seijo M.C., Escuredo O. & Rodríguez-Flores M.S., 2018. Characterization of Ziziphus lotus (jujube) honey produced in Algeria. J. Apic. Res., 57, 166-174, doi.org/10.1080/00218839.2017.1399663
Zerrouk S., Escuredo O., Rodríguez-Flores M.S. & Seijo M.C., 2021. Palynological characterisation of sedra honeys (Ziziphus lotus) produced in Algeria. Grana, 60(1), 69-80, doi.org/10.1080/00173134.2020.1770853
Zhang J. et al., 2020. Classification of the botanical and geographical origins of Chinese honey based on 1H NMR profile with chemometrics. Food Res. Int., 137(September), 109714, doi.org/10.1016/j.foodres.2020.109714
