


- Portada
- Volume 27 (2023)
- Numéro 1
- Folk taxonomical distinction between Irvingia gabonensis and I. wombolu (Irvingiaceae) in the Volta Forest region, West Africa


Vista(s): 2314 (36 ULiège)
Descargar(s): 132 (0 ULiège)

Folk taxonomical distinction between Irvingia gabonensis and I. wombolu (Irvingiaceae) in the Volta Forest region, West Africa

Documento adjunto(s)
Version PDF originaleRésumé
Distinction taxonomique traditionnelle entre Irvingia gabonensis and I. wombolu (Irvingiaceae) dans la région forestière de la Volta, Afrique de l’Ouest
Description du sujet. Irvingia gabonensis et I. wombolu produisent des amandes comestibles, avec des fruits respectivement sucrés et amers. Les chevauchements de caractères morphologiques et écologiques entretiennent un conflit taxonomique entre les deux espèces.
Objectifs. Cette étude évalue l’efficacité du système de classification endogène à séparer les deux espèces, I. gabonensis et I. wombolu, dans la région forestière de la Volta (Togo) où les deux espèces cohabitent.
Méthode. Cent-quatorze individus, des cinq principaux groupes socioculturels, ont été enquêtés afin d’identifier les critères de différentiation des deux espèces. Une reconnaissance en milieu naturel a été faite pour les arbres de I. gabonensis et de I. wombolu qui ont été évalués avec les facteurs écologiques localement utilisés dans la différentiation des deux espèces.
Résultats. Les résultats indiquent des variations au sein de chaque espèce. Mais ces variations sont non botaniques. Ainsi, la classification endogène était beaucoup plus basée sur le goût de la pulpe, ainsi que sur des critères écologiques. Les résidents locaux âgés considéraient le goût des fruits comme critère consistant de séparation des deux espèces. Seuls les jeunes hommes Akposso prennent également en compte l’écologie et la forme des feuilles pour la classification. Le test de reconnaissance en milieu naturel démontrait la consistance du système de classification endogène. Les paramètres dendrométriques de I. gabonensis dépendaient de l’environnement, alors que les systèmes d’utilisation des terres et la géomorphologie de sol influençaient significativement la présence de I. wombolu.
Conclusions. L’importance du goût des fruits et des saisons de fructification dans la séparation des deux espèces et l’échec des résidents âgés à utiliser les traits écologiques et botaniques dans la séparation démontre un contact récent des communautés de la région de la Volta avec ces taxa. Il est aussi plausible que la domestication amortit les différences entre I. gabonensis et I. wombolu.
Abstract
Description of the subject. Irvingia gabonensis and I. wombolu produce edible kernels, with their fruit pulp tasting sweet and bitter, respectively. The overlap of their morphological and ecological traits creates taxonomic uncertainty.
Objectives. This study assessed the efficiency of folk classification system for the separation between I. gabonensis and I. wombolu, in the Volta Forest region (Togo), where both species co-occur.
Method. Interviews were conducted with 114 respondents of the five main sociolinguistic groups in the Volta Forest region, in order to identify distinctive traits between both species. Field reconnaissance and confirmation test were used to identify I. gabonensis and I. wombolu trees, which were evaluated against ecological features in folk classification system.
Results. Variations existed within each species but were not botanically delimited. The folk classification system was mostly based on fruit pulp taste and some ecological traits. Old local residents only trusted fruit taste to separate these species. Only young Akposso men additionally considered ecology and leaf shape in classification. The reconnaissance survey confirmed the consistency of their folk classification system. Dendrometric parameters of I. gabonensis depended on environmental factors, while land use system and soil geomorphology significantly influenced the occurrence of I. wombolu.
Conclusions. Fruit pulp taste and flowering seasons were widely used to distinguish between species, while young men used ecological and botanical features. This demonstrates their recent contact with these taxa. It is also likely that domestication is narrowing differences between I. gabonensis and I. wombolu.
Tabla de contenidos
Received 17 January 2022, accepted 9 November 2022, available online 9 January 2023
This article is distributed under the terms and conditions of the CC-BY License (http://creativecommons.org/licenses/by/4.0)
1. Introduction
1Irvingia gabonensis and I. wombolu belong to the Irvingiaceae family, which occurs in West and Central Africa (Harris, 1996). These two indigenous fruit tree species have many botanical similarities and an overlapping natural range from the Casamance in southern Senegal to Angola, South-Sudan and Uganda (Harris, 1996; Baranga, 2007). This overlapping morphology and ecology is the source of taxonomic conflict between both species. Thus, I. gabonensis and I. wombolu are either considered as different varieties e.g. I. gabonensis var. gabonensis and I. wombolu var. excelsa (Mildbr.) Okafor, respectively (Okafor, 1975), or as separate species: Irvingia gabonensis (Aubry-Lecomte ex O'Rorke) Baill and I. wombolu Vermoesen, respectively (Harris, 1996; Lowe et al., 2000).
2Irvingia gabonensis and I. wombolu are respectively called sweet and bitter African bush mango trees (ABMTs) because of their sweet and extremely bitter/astringent mango-like fruits, respectively. The “Ogbono” or “Dika” soup, cooked with ground kernels of both types, belongs to the food culture in West and Central Africa (Vihotogbé et al., 2014). The commercialization of this non-timber forest product (NTFP) from sub-Sahara African countries generates significant income for local farmers, collectors and commercial actors (Leakey, 1999; Tabuna, 2000). Thus, ABMTs are priority agroforestry trees in West and central Africa because of the increasing demand and rising prices of the kernel (Leakey, 2012).
3Over the last three decades there has been a substantial increase in the domestication of indigenous fruits and nuts, especially in West Africa (Leakey et al., 2022) due to their potential importance in African agriculture and local economies. Nevertheless, there is a need to expand and enhance the domestication of species like the AMBTs with desirable traits (Ladipo, 1999). Unfortunately, the taxonomic complexity causes misidentification of elite trees for propagation. Consequently, intensive AMBT-based agricultural systems fail at reaching their potential to provide local livelihood improvement. Meanwhile, natural and anthropogenic factors contribute to a narrowing of the natural AMBT population size and diversity (Vihotogbé et al., 2013).
4Local Akébou, Akposso, Kabyè, Kotokoli and Ewé indigenous people hold valuable traditional ecological knowledge on surrounding biodiversity including the AMBT, which they classify based on natural and human-induced variations (Berlin et al., 1973). Better understanding of local plant names and biological variation through the local life classification system could provide valuable support for modern systematics (Costa-Neto, 1998) and so enhance the sustainable use and conservation of the local environment (Warren, 1992; Wilder et al., 2016).
5This study evaluates the diversity of the components in traditional folk classification systems of the Akébou, Akposso, Kabyè, Kotokoli and Ewé communities in Togo and their power to distinguish between I. gabonensis and I. wombolu. We used surveys to gather local residents’ knowledge regarding local names and the features that distinguish these two taxa in natural forests and in agroforestry systems. Then, the traditional plant systematics techniques of young Akposso were used to identify I. gabonensis and I. wombolu in the field and confirm the trees’ nature through the scoring of fruit pulp taste. This research approach was used to respond to two research questions:
6– Is there any difference in knowledge within and among sociolinguistic groups regarding the use of distinctive traits between I. gabonensis and I. wombolu?
7– How can folk classification be used for a successful in-field identification of I. gabonensis and I. wombolu?
2. Method
2.1. The study area
8This study was conducted in the district of Wawa in Togo’s western highlands in the Volta Forest region where I. gabonensis and I. wombolu co-occur in West Africa (Figure 1). It is a moist semi-deciduous forest zone (rain fall: 1,600 mm, temperature: 25 °C) around the Volta Lake from Eastern Ghana to Togo (Rödel & Agyei, 2003) where both the exploitation of natural forest bioresources by local residents and timber industries and their cash crop agriculture put a number of plant species at risk (Kuevi, 1981; Hillers et al., 2009). The seeds of I. gabonensis and I. wombolu are intensively collected in natural forests as well as in traditional agroforestry systems in the Volta Forest region and marketed around West Africa (Atato et al., 2010).
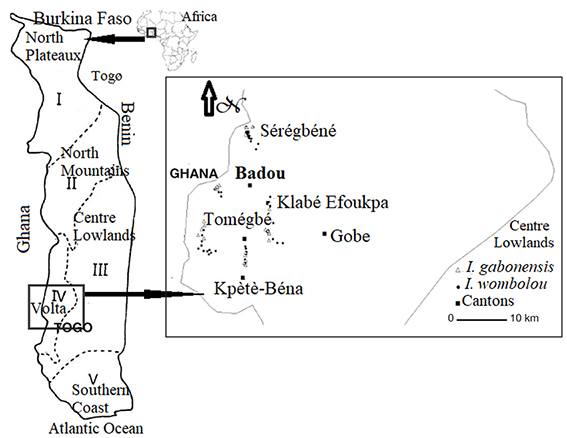 Figure 1. Location of the study area and geographical location of sampled trees in Togo — Situation géographique de la zone d’étude et des arbres échantillonnés au Togo.
Figure 1. Location of the study area and geographical location of sampled trees in Togo — Situation géographique de la zone d’étude et des arbres échantillonnés au Togo.
2.2. Components of traditional systematics system of I. gabonensis and I. wombolu
9Irvingia gabonensis and I. wombolu co-occur in the district of Wawa, here kernel samples were gathered at the main regional market of Badou-centre (Figure 1). Two villages in each canton, with at least one local collector of bush mango kernels, were randomly sampled to conduct surveys on the local systematics of ABMTs. Thus, a rapid appraisal was made with 40 randomly sampled respondents in the entire study area. The proportion (P) of positive answers was used to compute the numbers (n) of the individuals to be surveyed following Dagnelie (1988). In total, 114 respondents (9 per village) were randomly selected:
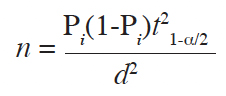
10with Pi: 95%, t1-α/2: 1.96 and a projected error (d) of 4%.
11In order to identify the components of folk classification of ABMTs, a structured questionnaire was used to record local sub-types of trees within I. gabonensis and I. wombolu populations, based on both botanical characteristics (i.e. leaves, fruits, and seeds), and local names associated to these sub-types. Moreover, botanical characters like phenology and non-botanical ones (e.g. soil, vegetation, land use system) and local names ascribed to these were recorded. Lastly, we recorded local perceptions of recent changes in I. gabonensis and I. wombolu (e.g., population size: increase, decrease or stability) including local perceptions of regeneration availability and the causes, diversity and impacts of threats to adult trees, along with the impacts of these trends on kernel prices.
2.3. Evaluating the effectiveness of ABMTs’ traditional systematics
12In order to assess accuracy of locally-used criteria to make field distinction between I. gabonensis and I. wombolu, and to increase detection probability of ABMTs in the low-density context of the Volta Forest ecoregion, large megaplot transects of 600 m x 60 m were used (Vihotogbé et al., 2014). At least 1 km distance separated consecutive megaplots, inside which secondary plots of 50 m x 30 m were established perpendicular to the swamps and stream belt to account for the high ecological variability within the area. In total, 96 secondary plots of 1,500 m2 across 12 megaplots were sampled in the study area, applying Dagnelie (1988) formula for:
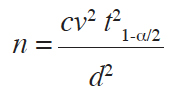
13with n: number on secondary plots; cv: 47.1%; t1-α/2: 2.064 and d: 10%.
14First, a multilinguistic group of 18 local experts, comprising three to four representatives of Akébou, Akposso, Kabyè, Kotokoli and Ewé was made and a reconnaissance survey was conducted in order to identify individual ABMTs (i.e., as belonging to the same plant known by the local name for either species) throughout every secondary plots. Individual ABMTs were regularly observed until fruiting period. In order to confirm field separation during the reconnaissance survey, five ripe fruits (already fallen from the tree) were collected from twenty I. gabonensis and I. wombolu trees randomly sampled in the secondary plots. Fruits were stored at room temperature during two days prior to a confirmation test. In March and June 2016, 42 independent volunteers with no prior knowledge on bitter versus sweet tasting bush mangoes, were recruited in Badou-Centre to taste and grade the fruit pulp from each sampled tree, using taste scoring system (Atangana et al., 2002). Thus, volunteers were trained to provide grades for bitterness degree to the samples: 0 (no bitter taste), 1 (very slightly bitter and eatable), 2 (mildly bitter and edible), 3 (bitter and not edible), and 4 (extremely bitter and even difficult to taste). Likewise, volunteers were also asked to provide similar grades for the sweetness degree of the samples: 0 (no sweet taste), 1 (very slightly sweet with almost water-tasting juice or with other dominant taste), 2 (mildly sweet with or without any other perceived taste), 3 (sweet with or without any other perceived taste), and 4 (extremely sweet with very slight and difficult to taste bitter taste). Finally, both fruit pulp types were similarly graded for sourness degree as for sweetness and bitterness. Here, bitter taste referred to quinine-like taste, while lemon/acid taste was considered as sourness.
15Second, occurrence, abundance and dendrometric data were collected in order to evaluate the accuracy of the traditional field identification process. Thus, within each secondary plot, presence/absence and total number of trees clearly identified as I. gabonensis versus I. wombolu after reconnaissance test, were recorded. Individual trees were measured for their diameter at breast height (DBH) and individual secondary plots were characterized along with environmental factors locally indicated as governing field distinction between I. gabonensis and I. wombolu: elevation at the center of each secondary plot using GPS, soil geomorphology (e.g. plateau, hill, slope, temporarily flooded, valley and watershed), mean slope value (center of each secondary plot using the angle described by soil inclination with the vertical), presence/absence (1/0) of permanent swamp, type of land use system (natural forest, plantation, or home garden) and drainage level (highly drained or not drained). Moreover, percentage of ground grasses and tree cover, explaining the degree of habitat savannization, due to anthropogenic changes and natural conditions in the Volta Forest ecoregion (Kasei, 2009) were estimated and cover coefficient was attributed to individual plots (0-1% = 0.5; 1-5% = 3; 5-25% = 15: 25-50% = 37.5; 50-75% = 62.5 and 75-100% = 87.5) (Braun-Blanquet, 1932). The distance separating individual trees to the nearest stream was measured and four soil samples at 30 to 40 cm depth around each tree as well as in the middle of plots were taken and mixed for rapid on-farm granulometric analysis to determine clay, silt and sand contents (Robert & Saucier, 1988).
2.4. Data analysis
16Traditional systematics of I. gabonensis and I. wombolu. Three sub-groups of respondents were defined within each sociolinguistic group based on the age profile: young (< 30 years), adults (30 > and < 60 years), and elderly (> 60 years) (Teka & Vogt, 2010). Respondents were also professionally categorized: wood sawyers, wood loggers, wood businessmen, wood marketers, local farmers, and others. Therefore, for each sub-group and within each sociolinguistic group we calculated the percentage of respondents who mentioned each character as significantly discriminative of I. gabonensis and I. wombolu. In order to associate discriminating characters with sub-groups, a Principal Component Analysis (PCA) was performed using the package FactoMineR (https://r-project.org/), on the frequency-based data matrix, using R software (version 3.2.2). Professional categorization was used in PCA as an illustrative variable (Santos, 2015).
17Using descriptive statistics, local perceptions of I. gabonensis and I. wombolu population dynamics were assessed based on locals’ responses regarding increase, decrease or stability in population size, regeneration availability, diversity of threats and the effect on variation in kernel prices.
18Differentiation of I. gabonensis and I. wombolu. Using scores provided by respondents in the sensorial analysis, tree-to-tree variation of bitterness, sweetness and sourness was assessed, using descriptive statistics. Since variation in the levels of sweet and bitter tastes was detected for I. gabonensis, a one way-ANOVA was carried out on bitter taste scores against those of sweet taste, as revealed by local tasters, in order to identify the dominant taste of the fruit pulp of this species.
19For each plot, the density (N) of adult trees (individuals/hectare) was calculated for I. gabonensis and I. wombolu. The mean basal area (G) and quadratic diameter (D) were evaluated as follow:
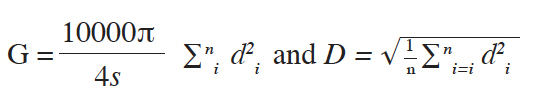
20di being the diameter of ith randomly sampled tree in the plot of area s = 1,500 m².
21In order to evaluate the impact of local environmental factors on the abundance and dendrometric performances of I. gabonensis and I. wombolu, we used the Generalized Linear Model to apply multiple linear regressions on the density, mean basal area and quadratic diameter data against altitude, slope, ground grasses and trees covers, soil clay, silts and sand contents and mean distance from the nearest stream. This analysis was complemented with a binary logistic regression performed on land use system, presence of flood, geomorphology and drainage against presence versus absence of I. gabonensis and I. wombolu in order to evaluate how these categorical factors influence the occurrence probability of ABMTs in the Volta Forest region. Finally, one-way analysis of variance (ANOVA) was applied on density, mean basal area and quadratic diameter data to evaluate the difference between the two types of ABMTs. Before the analysis, we checked normality (Ryan-Joiner test) and homoscedasticity (Levene test) of the variances (Glèlè Kakaï et al., 2006) and applied square-root transformation on density data and log-transformed the basal area and quadratic diameter (O’Hara & Kotze, 2010).
3. Results
3.1. Local application of the components of traditional distinction system of ABMTs
22No traditional classification system, based on any botanical or organoleptic character was identified for I. gabonensis. However, all respondents recognized remarkable tree-to-tree variations in tree shape, fruit juice/fiber content, fruit taste, size, kernel color, sliminess, and aroma. Even though these variations guide local residents’ food choice, corresponding trees have never been botanically classified or delimited, in order to be locally named.
23Seventy-eight percents of the respondents were unable to botanically distinguish variation in I. gabonensis because materials with identical morphological vegetative characteristics often produce fruits with a wide range of fruit morphology and taste, even in the same ecological conditions.
24Likewise, for I. wombolu, only 8% of local residents distinguished between two main fruit shapes: spheroid and oblong. Within these types, they distinguished different sized fruits, regardless of the mother trees’ age and ecological environment. However, no vernacular name distinguished such morphological variation in the field.
25Irvingia gabonensis was called “Atokliko” in the Volta Forest region, while the local residents of Akposso called I. wombolu “Aakpa” and this name was known by 33% of Akébou and 2% of Kotokoli. In general, there was no diversity in local names for I. gabonensis and I. wombolu corresponding to the sociocultural diversity. Instead, the respondents for ABMTs in the study area used the terms “bitter and never cultivated ABMTs” versus “sweet and cultivated ABMTs” for I. gabonensis and I. wombolu, respectively. Folk classification combined leaf shape with non-botanical characters of fruits and trees, as well as the ecology and phenology to distinguish between both ABMT taxa in the field. All the respondents acknowledged that fruits of I. wombolu are extremely bitter, harsh, and thus completely inedible. In contrast, those of I. gabonensis are edible because they are sweet and never astringent, even though a slightly bitter aftertaste is often noticeable. According to young Akposso men, given the effective discriminative power of this character, its evaluation should be focused on the detection of a minimum sweet taste in the pulp, which never overlaps between both species, this being the most consistent trait used to separate I. gabonensis and I. wombolu.
26Ecologically, 84.1% of the respondents asserted that although both species co-occur on many soil types in the Volta Forest region, I. wombolu is a forest tree species while I. gabonensis is mostly found in traditional agroforestry systems. Local residents (52%) asserted that I. wombolu is commonly found in swampy areas and stream banks.
27In combination with fruit taste, tree size (total height and DBH) was also an important character used by 27.3% of the respondents, to distinguish between both species. According to folk classification, mature trees of I. gabonensis are smaller and shorter than mature individuals of I. wombolu. Leaf shape was also combined with fruit taste and ecology by 19.3% of the respondents to distinguish the two taxa. In general, leaves of I. gabonensis were said to be longer and narrower, and more elliptical than those of I. wombolu, which were more obovate than those of I. gabonensis.
28Phenologically, 100% of local residents were unanimous that the fruiting season of I. wombolu is the dry season (January, February, March), whereas I. gabonensis was always characterized by a rainy season fruiting (May-September).
29The first two components of the PCA to distinguish the two species explained 74.7% of the total variation within the dataset (Table 1). The first component (49.4%) was that of fruit pulp taste with land use system (presence in flooded areas and along streams), whereas the second component (25.4%) was tree size (Figure 2). It is also apparent that four groups can be identified, which correspond to clearly distinct folk classification systems (Figure 2). The first group was composed of young Akposso men, who considered differences in fruit taste, ecology (land use systems, presence of swamps and stream) and leaf shape to distinguish I. wombolu from I. gabonensis (G1, Figure 2). In contrast, adult Akebu men and young Ewé men (G2, Figure 2) mostly focused on ecological characteristics related to wet sites to distinguish the taxa. These first two groups did not acknowledge dendrometric properties as consistent distinctive characters between I. gabonensis and I. wombolu whereas dendrometric characteristics along with land use systems were a very important part of folk classification for the third group - the young Kabiyè men, adult Akebu, Kabiyè women, and adult Akposso men (G3, Figure 2). The fourth group (G4, Figure 2) was the most age-heterogeneous, including elderly men of all sociocultural groups. Members of this group recognized differences in fruit taste but emphasized overlap of every character that composed folk classification to distinguish I. wombolu from I. Gabonensis.
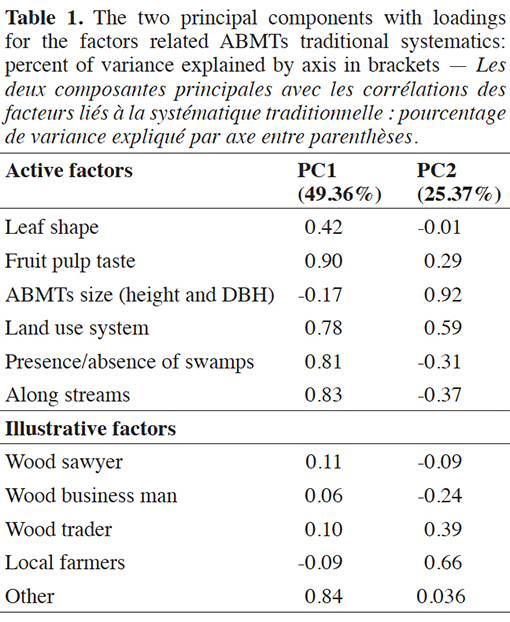
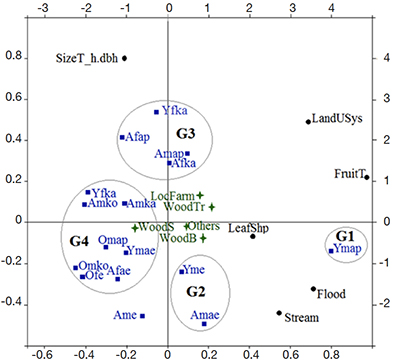
Figure 2. Results of the Principal Component Analysis showing ethnic sub-groups with similar traditional taxonomic system for Irvingia gabonensis and Irvingia wombolu — Résultats de l’Analyse en Composantes Principales montrant les sous-groupes ethniques avec le même système taxonomique traditionnel pour Irvingia gabonensis et Irvingia wombolu.
A: adults — adultes; Y: young — jeunes; O: old — vieux; f: female — femmes; m: male — hommes; e: Ewe; ae: Akebu; ap: Akposso; ko: Kotokoli; Ka: Kabiyè.
3.2. Local perception of population dynamics and threats on kernels price
30Local residents reported a rapid decrease of the wild I. wombolu population size and an increase in the planted I. gabonensis population (68% and 66%, respectively). However only 6.8% and 16% had the inverse perception for I. wombolu and I. gabonensis, respectively. In addition, 20.4% and 18% of the respondents acknowledged a constant population size for I. wombolu and I. gabonensis, respectively.
31Irvingia wombolu trees were more intensively exploited (70.5%) than I. gabonensis (16%) being felled for timber, according to local residents. Extension of cultivation fields, for subsistence agriculture as well as for cash crops like cocoa and coffee, were the second most important factor threatening adult individuals of both taxa (63.6% versus 68.2% for I. gabonensis and I. wombolu, respectively). Intensive collection of I. wombolu and I. gabonensis fruits for the local kernel trade threatened regeneration according to 97.7% and 95.5% of the respondents, respectively. One hundred percent of the respondents considered the elimination of saplings during the preparation of agricultural fields to be a severe threat for both taxa (Figure 3).
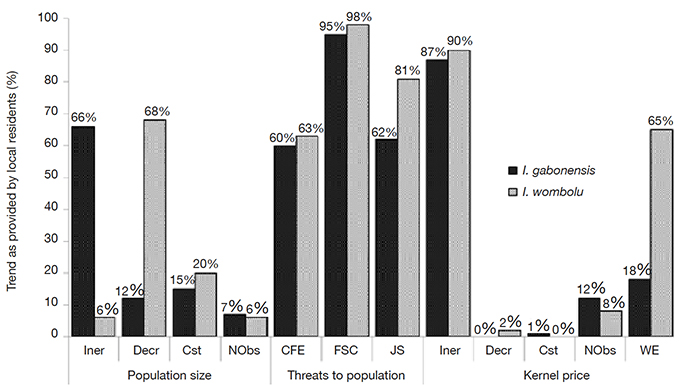 Figure 3. Recent perception of local residents regarding differences of fluctuations in population size, diversity and level of threats and their impacts on Irvingia gabonensis and I. wombolu kernel prices — Perception récente des communautés locales à propos des différences de fluctuation de la taille des populations, de la diversité et des niveaux de menaces et leurs impacts sur les amandes de Irvingia gabonensis et I. wombolu.
Figure 3. Recent perception of local residents regarding differences of fluctuations in population size, diversity and level of threats and their impacts on Irvingia gabonensis and I. wombolu kernel prices — Perception récente des communautés locales à propos des différences de fluctuation de la taille des populations, de la diversité et des niveaux de menaces et leurs impacts sur les amandes de Irvingia gabonensis et I. wombolu.
Incr: increase — augmentation; Decr: decrease — diminution; Cst: constant — constant; Nobs: no observation — pas d’observation;WE: wood exploitation — exploitation de bois; CFE: cultivation field extension — agriculture extensive; FSC: fruit and seed collection — collection de fruits et graines; JS: juvenile suppression — éradication des juvéniles.
32Almost 90% of respondents mentioned an increase in kernel price as a direct impact of threats to Irvingia populations. Moreover, despite the increase of I. gabonensis population size, its kernel price was still increasing (Figure 3).
3.3. Diversity of fruit pulp taste and difference in their spatial distribution
33The fruit pulp taste grading indicated no diversity of taste for I. wombolu fruit pulp, which was consistently reported as extremely bitter and difficult to taste (grade 4) by 100% of the tasters for all sample, with a total zero sweetness and sourness. In contrast, different degrees of sweetness and bitterness plus total absence of sour taste were recorded for I. gabonensis fruit pulps (Figure 4b). Thus, sweetness scores ranged between 2 and 4 and bitterness between 0 and 1 for I. gabonensis. The one-way ANOVA revealed that the pulps of I. gabonensis are sweeter than bitter (p < 0,0001; Figure 4). Unfortunately, only 15% of I. gabonensis trees produced totally sweet fruits (with sweetness grade = 4, bitterness grade = 0, and sourness grade = 0).
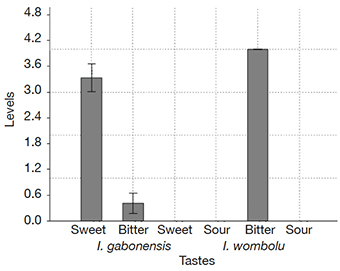 Figure 4. Mean sweetness, bitterness, and sourness scores in the fruit pulp of Irvingia gabonensis and I. wombolu — Moyenne des scores des goûts sucré, amer et acide dans les pulpes de Irvingia gabonensis et I. wombolu.
Figure 4. Mean sweetness, bitterness, and sourness scores in the fruit pulp of Irvingia gabonensis and I. wombolu — Moyenne des scores des goûts sucré, amer et acide dans les pulpes de Irvingia gabonensis et I. wombolu.
34Results of multiple linear regressions indicated that in the Volta Forest region environmental factors predict density, basal area and quadratic diameter of I. gabonensis and I. wombolu (Table 2; Figure 5). Variation in the distance to the nearest stream and soil clay content significantly influenced the density of I. gabonensis; while only tree cover significantly defined differences in its basal area and soil silt content significantly impacted on quadratic diameter of this taxon (Table 2). In contrast, densities of I. gabonensis significantly decreased with increasing distance from the nearest stream. High densities of I. gabonensis were predicted at around 150-350 m from streams, while an increase of clay content in the soil also increased the density of this species in this study. Moreover, an increase of tree cover significantly reduced I. gabonensis basal area. Likewise, stem diameter of I. gabonensis significantly decreased with increasing density of tree cover. On the other hand, soil silt content was significantly linked with low values of this dendrometric parameter (Table 2; Figure 5). Regarding I. wombolu, none of the environmental factors accountable in the traditional systematics significantly explained density, basal area and quadratic diameter in this forest.
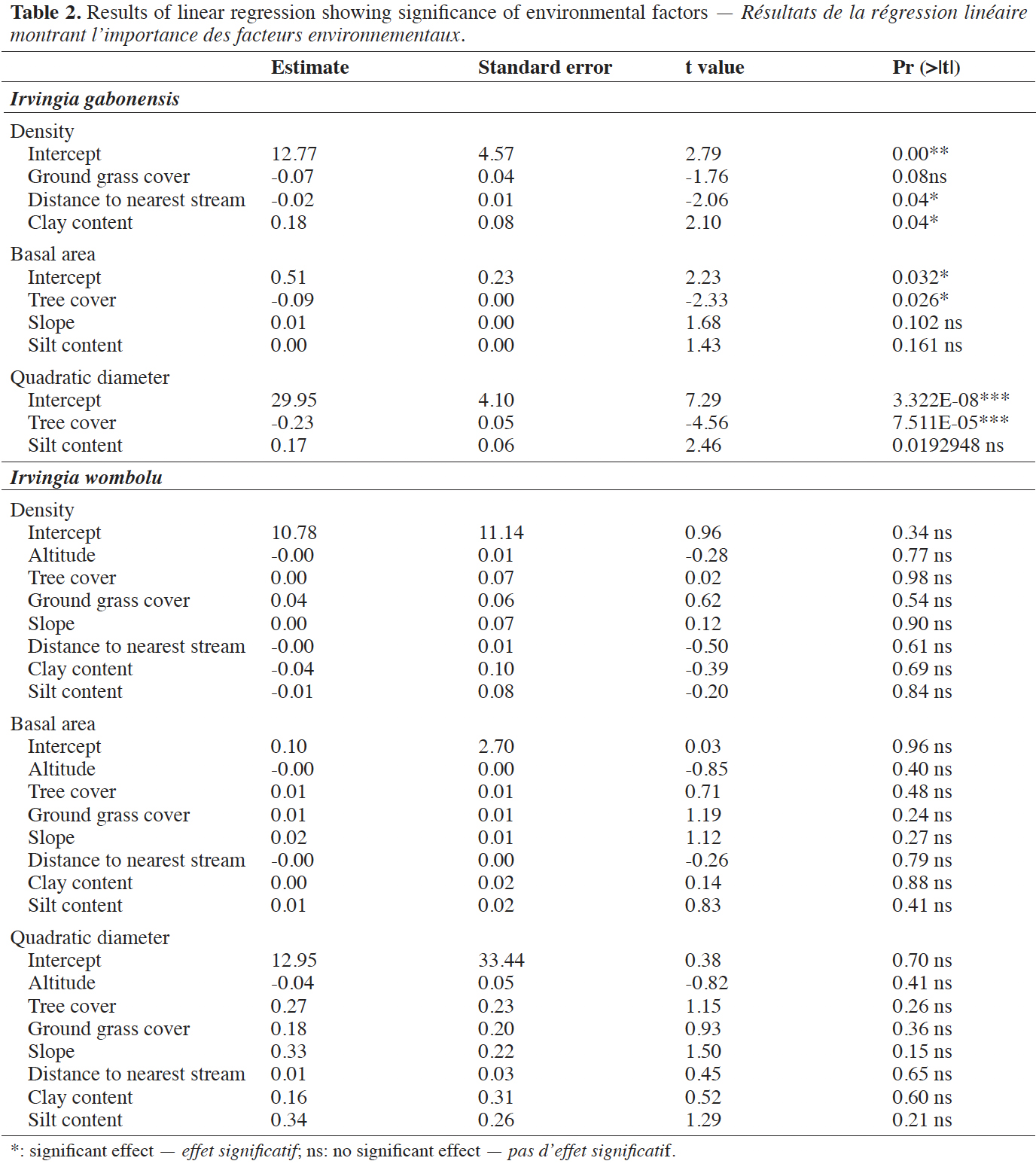
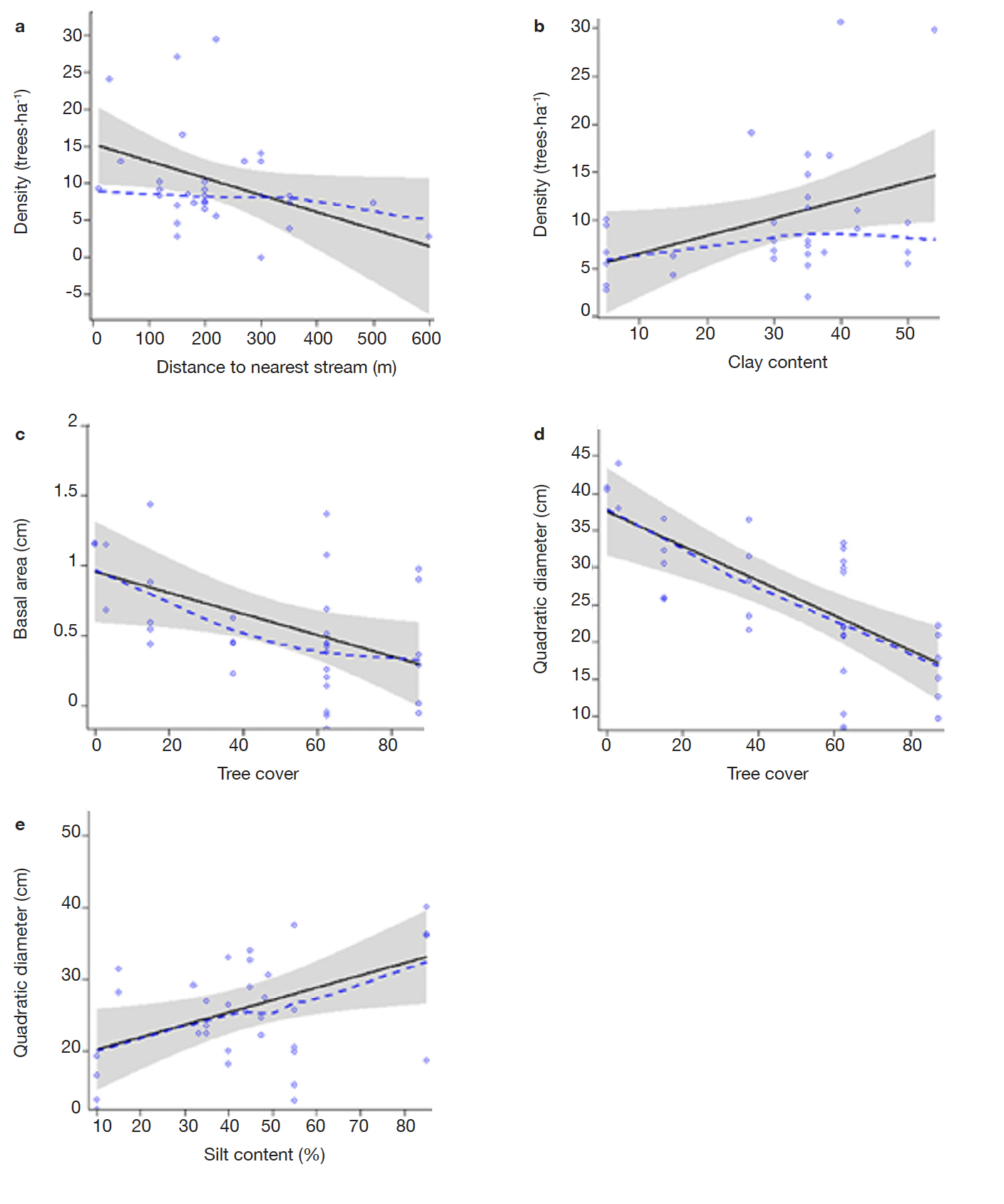
Figure 5. Results of multiple linear regression assessing predictability of Irvingia gabonensis’ density and dendrometric parameters with environmental factors: a. density against distance to the nearest stream, b. density against soil clay content, c. basal area against soil tree cover, d. quadratic diameter against soil tree cover, e. quadratic diameter against soil silt content. No environmental factor significantly influences density and dendrometric characteristics of I. wombolu — Résultats de la régression linéaire multiple évaluant la prédiction de la densité de Irvingia gabonensis et les paramètres dendrométriques avec les facteurs environnementaux : a. densité contre la distance au cours d’eau le plus proche, b. densité contre la teneur en argile du sol, c. surface basale contre la couverture arborée du sol, d. diamètre quadratique contre la couverture arborée du sol, e. diamètre quadratique contre la teneur en limon du sol. Aucun facteur environnemental n’influence la densité et les caractéristiques dendrométriques de I. wombolu.
35In the Volta Forest region, the logistic regression indicated that occurrence of I. gabonensis varied with the land use system and the occurrence of low-lying land susceptible to flooding, while land use system and slope explained that of I. wombolu (Table 3). Thus, I. gabonensis had higher occurrence probability in human made forest-like agroforestry systems (plantations) and in non-flooded areas, while I. wombolu had a greater probability of occurrence in natural forests on soils with low and moderately steep slopes.
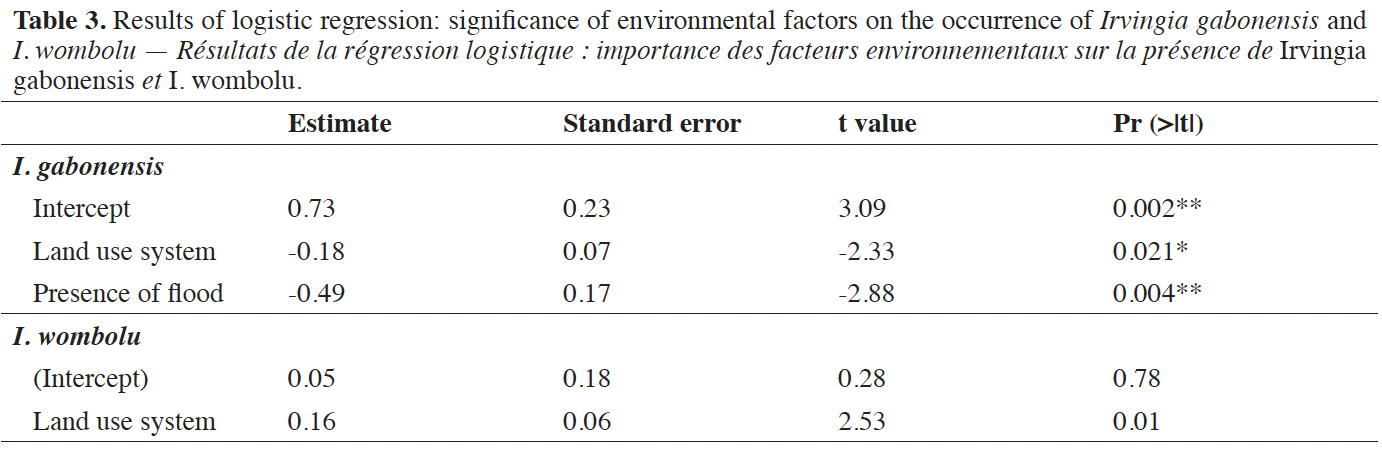
36Results from the ANOVA test (Table 4) indicated significant difference between I. wombolu and I. gabonensis, regarding their mean density (P = 0.002) and mean quadratic diameter (P = 3.4 E-09). However, mean basal area was not significantly different between the two taxa.

4. Discussion
4.1. Variation in the traditional taxonomy of ABMTs
37Results suggest that local people in the Volta Forest region have poor knowledge, regarding classification within I. wombolu and I. gabonensis, based on morphological variability. This is in line with a low level of ethnobotanical knowledge related to traditional uses of these resources (Vihotogbé et al., 2014) and may be explained by the relatively recent contact of local residents with I. gabonensis in the study area and by rapid changes in the landscape and natural resources with inadequate management strategies. Indeed, only I. wombolu is demonstrated to be native to this research area and is abundant in natural ecosystems in the Volta Forest area, which does not belong to the ecological niche of I. gabonensis (Vihotogbé et al., 2019). This species is found only in agroforestry systems, with younger populations (Vihotogbé et al., 2014). However, even this limited ecological knowledge is being lost due to rapid changes in the landscape and the lack of adequate management strategies for natural resources (Nolan, 2002). Nevertheless, it is clear that young Akposso residents rely on a combination of botanical and non-botanical traits to differentiate I. gabonensis from I. wombolu in the Volta Forest region, and their classification system based on the taste of fruit pulp was found to be effective during field reconnaissance survey, based on fruit pulp taste. This demonstrates that members of this subgroup have advanced expertise in taxonomy because leaf morphology remains central to plant taxonomy (Viscosi & Cardini, 2012), and biodiversity conservation (Nolan, 2002; Peterson, 2006). The integration of ecological and phenological features in their local systematics system reflects modern approaches and demonstrates the need for more complex biological evaluation systems for closely related plant taxa (Mustapha, 2009; Olango et al., 2014). However, limited botanical traits are used locally and bush mango pulp taste remains central in the traditional systematics of ABMTs. This reflects the influence of food culture on traditional knowledge related to food resources (Otieno et al., 2015). Therefore, ascribing vernacular names that describe bitter versus sweet properties of the pulp supports the central role of descriptive naming process related to traditional use in plant species (Otieno et al., 2015). These description-based names also make sense in the classification process of kernels, considering that kernels have greater cultural importance and higher economic value for their use as food thickening agents in local cuisine (Jamnadass et al., 2011). However, ecological, phenological and dendrometric parameters also have a role in local systematics and are used to guide seed collection for kernel extraction. Phenology of plant species and particularly that of ABMTs is ecology-dependant and subjected to natural and anthropogenic factors (Pau et al., 2011; Vihotogbé et al., 2014). Moreover, tree-to-tree dendrometric variations (total height and DBH) depend on climatic, soil and local management systems as well as on age and taxonomic group (Brien et al., 1995; Lukaszkiewicz & Kosmala, 2008; Vanclay, 2009). Consequently, descriptive ecology and phenology data have limited power in the taxonomy of these ABMT taxa in which large-scale co-occurrence exists for I. gabonensis or I. wombolu throughout their entire geographical range (Harris, 1996), as well as different cultivation and occurrence histories in the Volta Forest region (Vihotogbé et al., 2014).
38Our data showed four clearly different groups of respondents among our native local residents in the Volta Forest region (Kouevi, 1970) (Figure 2). The ecological and phenological knowledge of the Akebu and Akposso socio-cultural groups that share environmental conditions and understanding was the most reliable to distinguish between species taxonomically.
39It is also reported that field distinction between I. gabonensis and I. wombolu is mostly based on fruit pulp taste, botanical features such as leaf shape and soil drainage level and water availability (Harris, 1996; Orwa et al., 2009). Young Akposso men and adult Akebu men were found to have consistent knowledge of the use of these criteria in distinguishing I. gabonensis from I. wombolu on the field. Apart from the Ewé local residents, all other interviewed communities are primarily involved in either the collection and marketing of bush mango kernels, or in timber business. It is therefore likely that the inability of many sub-groups (most importantly the old men) to distinguish between species using local systematics is rooted in the unclear morphological segregation of these co-occurring species (Vihotogbé et al., 2013). Nevertheless, it is likely that local residents know the location and fruiting time of every single ABMT, because of the limited population size and difficulties in observing detailed distinctive botanical characters on tall and mature I. wombolu trees. We also assume that older residents in the area lack this knowledge because of the recent introduction of cultivation of I. gabonensis in the Volta Forest area.
40African mangoes kernels are becoming increasingly expensive in the area and in surrounding areas. This is mainly due to the declining population of I. wombolu. In Central Africa, it has been reported that there is a correlation between population size and kernel market value (Anegbeh et al., 2003; Vihotogbé et al., 2008) as kernels have traditionally been collected from natural forests resulting in human-induced threats to the species (Lowe et al., 2000). Therefore, absence of cultivation of I. wombolu demonstrates that higher prices of its kernels are not yet a sufficiently motivating factor in a major cash crop (cocoa, bananas, ginger, coffee, etc.) producing area like the Volta Forest region.
4.2. Ecological distinction between I. gabonensis and I. wombolu
41The occurrence of I. gabonensis in traditional agroforestry systems in the Volta Forest region is typically in areas with red ferralitic clay soil where it is limited by water-logging as reported for coastal areas in West and Central Africa (FAO–UNESCO, 1977; Kang & Tripath, 1992). In contrast, I. wombolu grows well on both swampy and well drained soils with a wide range of granulometry, demonstrating its greater ecological adaptability (Turrill, 1964). Unfortunately, dedication of productive land to cash crops (cocoa, coffee, bananas, etc.) cultivation directly competes with Irvingia species in the Volta Forest region. Moreover, local communities’ food culture already integrates the sweet African mango pulp, but not yet totally the kernel-based slimy sauce (Vihotogbé et al., 2014), hampering initiatives for I. wombolu.
42This study establishes evidence that components of ABMTs’ local systematics may be valuable to distinguish I. gabonensis from I. wombolu as different fruit pulp tastes seem to be weakly correlated with ecology, while the other components of traditional systematics also predict little differences between these two taxa. Thus, folk knowledge does not support the suggestion that the taste of I. gabonensis and I. wombolu fruits distinguish them as ecotypes of a single Irvingia species. Further update of genetic studies is therefore required to understand the taxonomy of these species.
5. Conclusions: implications and limits of the study
43The present study reveals:
44– that significant variation in fruit pulp taste does exist between these species and between individual trees of I. gabonensis;
45– that fruit pulp taste and fruiting seasons were the characters unanimously recognized in folk systematics as distinct between these species. This brings out the importance of folk classification in sustaining biodiversity and the importance of the transmission of traditional ecological knowledge through the generations. Interestingly, however, only young men Akposso used ecology and leaf shape to distinguish between I. gabonensis and I. wombolu;
46– dendrometric parameters of I. gabonensis were significantly influenced by distance to the nearest stream, soil clay and silt content, and by landscape tree cover whereas none of those factors influenced the dendrometric parameters of I. wombolu. This knowledge may be important in future tree planting and agroforestry projects to produce kernels for domestic use and local trade.
47The limitations of the present study include:
48– the focus on fruit pulp taste rather than on kernel traits;
49– the risks of potentially poor or inaccurate translation and enumerator error when working on folk knowledge.
Acknowledgements
50We are grateful to the International Foundation for Science (IFS / Grant No: D/4672-1 to the first author) and to the donors for funding this research. Zaka Tagbekou and Nassirou from Kpete-Bena, in the Volta Forest region, assisted in field works and help recruit tasters in Badou. We thank them here, as well as the locals who freely participated to the field works in Badou, Tomegbe, Kpete-Bena, Kpete-Maflo and Kedjebi.
Bibliographie
Anegbeh P.O. et al., 2003. Domestication of Irvingia gabonensis: 3. Phenotypic variation of fruits and kernels in a Nigerian village. Agrofor. Syst., 58(3), 213-218, doi.org/10.1023/A:1026094021408
Atangana A.R. et al., 2002. Domestication of Irvingia gabonensis: 2. The selection of multiple traits for potential cultivars from Cameroun and Nigeria. Agrofor. Syst., 55(3), 221-229, doi.org/10.1023/A:1020584823505
Atato A. et al., 2010. Diversity of edible wild fruits species of Togo. Global Sci. Books, 4(1),1-9.
Baranga D., 2007. Observations on resource use in Mabira Forest Reserve, Uganda. Afr. J. Ecol., 45(1), 2-6, doi.org/10.1111/j.1365-2028.2007.00729.x
Berlin B., Breedlove D.E. & Raven P.H., 1973. General principles of classification and nomenclature in folk biology. Am. Anthropol., 75, 214-242, doi.org/10.1525/AA.1973.75.1.02A00140
Braun-Blanquet J., 1932. Plant sociology. New York, NY, USA: McGraw-Hill.
Brien S.T. et al., 1995. Diameter, height, crown, and age relationships in eight neotropical tree species. Ecology, 76(6), 1926-1939, doi.org/10.2307/1940724
Costa Neto E.M., 1998. Folk taxonomy and cultural significance of “abeia” (Insecta, Hymenptera) to the Pankararé, Northeastern Bahia State, Brazil. J. Ethnobiol., 18(1), 1-13.
Dagnelie P., 1988. Statistiques théoriques et appliquées. Bruxelles: De Boeck.
Glèlè Kakaï R., Sodjinou E. & Fonton H.N., 2006. Conditions d’application des méthodes statistiques paramétriques. Porto-Novo : Bibliothèque Nationale du Bénin.
Harris D.J., 1996. A revision of the Irvingiaceae in Africa. Bull. Jardin Bot. Belg., 65(1-2),143-196, doi.org/10.2307/3668184
Hillers A. et al., 2009. Assessment of the amphibians in the forests of southern Ghana and western Togo. Zoosyst. Evol., 85(1), 127-141, doi.org/10.1002/zoos.200800019
Jamnadass R.H. et al., 2011. Improving livelihoods and nutrition in sub-Saharan Africa through the promotion of indigenous and exotic fruit production in smallholders’ agroforestry systems: a review. Int. For. Rev., 13(3), 338-354, doi.org/10.1505/146554811798293836
Kang B.T. & Tripath B., 1992. Technical paper 1: soil classification and characterization. In: Tripath B.R. & Psychas P.J., eds. Alley farming research network for Africa, The AFNETA alley farming training manual. Ibadan, Nigeria: International Institute of Tropical Agriculture.
Kasei R.A., 2009. Modelling impacts of climate change on water resources in the Volta Basin, West Africa. PhD thesis: University of Bonn (Germany).
Kouevi A., 1970. Traditions, histoire et organisation de la cité chez les Akposso (cas de Uviu, de Emla et de Uzitse). Mémoire : Office de la Recherche Scientifique et Technique Outre-Mer.
Kuevi D., 1981. Ethnies et langues. Atlas Jeune Afrique– Togo. Paris : Les Éditions j.a.
Ladipo D.O., 1999. The development of quality control standards for Ogbono (Irvingia gabonensis and Irvingia wombolu) kernels; efforts towards encouraging organized and further international trade in a non-wood forest product of West and Central Africa. In: Sunderland T.C.H., Clark L.E. & Vantomme P, eds. Non-wood forest products of Central Africa: current research issues and prospects for conservation and development. Roma: FAO, 45-250.
Leakey R.R.B., 1999. Potential for novel food products from agroforestry trees. Food Chem., 64, 1-14, doi.org/10.1016/S0308-8146(98)00072-7
Leakey R.R.B., 2012. Living with the trees of life: towards the transformation of tropical agriculture. 1st ed. Wallingford, UK: CABI.
Leakey R.R.B. et al., 2022. The future of food: domestication and commercialization of indigenous food crops in Africa over the third decade (2012–2021). Sustainability, 14(4), 2355, doi.org/10.3390/su14042355
Lowe A.J.A., Gillies C.M., Wilson J. & Dawson I.K., 2000. Conservation genetics of bush mango from central/west Africa: implications from random amplified polymorphic DNA analysis. Mol. Ecol., 9, 831-841, doi.org/10.1046/j.1365-294x.2000.00936.x
Lukaszkiewicz J. & Kosmala M., 2008. Determining the age of streetside trees with diameter at breast height-based multifactorial model. Arboriculture Urban For., 34(3), 137-143, doi.org/10.48044/jauf.2008.018.
Mustapha O.T., 2009. Principles and practice of systematics. Int. J. Adv. Biotechnol. Res., 1(2), 126-133, doi.org/10.7591/cornell/9781501752773.003.0001
Nolan J.M., 2002. Wild plant classification in Little Dixie: variation in a regional culture. J. Ecol. Anthropol., 6, 68-80, doi.org/10.5038/2162-4593.6.1.5
O’Hara R. & Kotze J.D., 2010. Do not log-transform count data. Methods in ecology and evolution, 1, 118-122, doi.org/10.1111/j.2041-210X.2010.00021.x
Okafor J.C., 1975. Varietal delimitation in Irvingia gabonensis (Irvingiaceae). Bull. Jardin Bot. Nat. Belg., 45, 211-221, doi.org/10.2307/3667601
Olango T.M., Tesfaye B., Catellani M. & Pè M.E., 2014. Indigenous knowledge, use and on-farm management of enset (Ensete ventricosum (Welw.) Cheesman) diversity in Wolaita, Southern Ethiopia. J. Ethnobiol. Ethnomed., 10(41), 1-18, doi.org/10.1186/1746-4269-10-41
Orwa C.A., Mutua-Kindt R., Jamnadass R. & Anthony S., 2009. Agroforestree database: a tree reference and selection guide version 4.0, https://www.worldagroforestry.org/publication/agroforestree-database-tree-reference-and-selection-guide-version-40, (22/11/22).
Otieno J. et al., 2015. Vernacular dominance in folk taxonomy: a case study of ethnospecies in medicinal plant trade in Tanzania. J. Ethnobiol. Ethnomed., 11(10), 1-15, doi.org/10.1186/1746-4269-11-10
Pau S. et al., 2011. Predicting phenology by integrating ecology, evolution and climate science. Global Change Biol., 17, 3633-3643, doi.org/10.1111/j.1365-2486.2011.02515.x
Peterson A.T., 2006. Taxonomy is important in conservation: a preliminary reassessment of Philippine species-level bird taxonomy. Bird Consulting Int., 16(2), 155-173, doi.org/10.1017/S0959270906000256
Robert D. & Saucier J.P., 1987. Cartographie écologique à l'échelle 1:20 000. Service de l'inventaire forestier, Ministère de l'Énergie et des Ressources du Québec.
Rödel M.O. & Agyei A.C., 2003. Amphibians of the Togo Volta highlands, eastern Ghana. Salamandra, 39, 207-234.
Stone A. et al., 2000. Évaluation des échanges des produits forestiers non ligneux entre l’Afrique subsaharienne et l’Europe. FAO, Bureau Régional pour l’Afrique.
Teka O. & Vogt J., 2010. Social perception of natural risks by local residents in developing countries, the example of the coastal area of Benin. Social Sci. J., 47, 215-224, doi.org/10.1016/j.soscij.2009.07.005
Turrill W.B., 1964. The ecotype concept. A consideration with appreciation and criticism, especially of recent trends. New Phytol., 45(1), 34-43, doi.org/10.1111/j.1469-8137.1946.tb05044.x
Vanclay J.K., 2009. Tree diameter, height and stocking in even-aged forests. Ann For. Sci., 66(702), 17, doi.org/10.1051/forest/2009063
Vihotogbé R. et al., 2008. Ethnobotany and endogenous conservation of Irvingia gabonensis (Aubry-Lecomte) Baill. in traditional agroforestry systems in Benin. Indilinga, 6(2), 196-209, doi.org/10.4314/indilinga.v6i2.26428
Vihotogbé R., Berg van Den R.G. & Sosef M.S.M., 2013. Morphological characterization of African bush mango trees (Irvingia species) in the Dahomey Gap (West Africa). Genet. Resour. Crop Evol., 60, 1597-1614, doi.org/10.1007/s10722-013-9969-0
Vihotogbé R. et al., 2019. Ecological niche information supports taxonomic delimitation of Irvingia gabonensis and I. wombolu (Irvingiaceae). South Afr. J. Bot., 127, 35-42, doi.org/10.1016/j.sajb.2019.08.025
Vihotogbé R. et al., 2014. Impacts of the diversity of traditional uses and potential economic value on food tree species conservation status: case study of African bush mango trees (Irvingiaceae) in the Dahomey Gap (West Africa). Plant Ecol. Evol., 147(1), 109-125, doi.org/10.5091/plecevo.2014.789
Viscosi V. & Cardini A., 2012. Leaf morphology, taxonomy and geometric morphometrics: a simplified protocol for beginners. PLoS ONE, 7(3), doi.org/10.1371/annotation/bc347abe-8d03-4553-8754-83f41a9d51ae
Warren D.M., 1992. Indigenous knowledge, biodiversity conservation and development. Keynote address: International Conference on Conservation of Biodiversity in Africa: Local Initiatives and Institutional Roles, 30 August-3 September, Nairobi, Kenya.
Wilder B.T., O’meara C., Monti L. & Nabhan G.P., 2016. The importance of indigenous knowledge in curbing the loss of language and biodiversity. BioScience, 66(6), 499-509, doi.org/10.1093/biosci/biw026
