


- Accueil
- Volume 24 (2020)
- Numéro 1
- May passive dispersal of fungal enemies with native substrates in olive nurseries help to control phytonematodes?


Visualisation(s): 2349 (41 ULiège)
Téléchargement(s): 468 (10 ULiège)

May passive dispersal of fungal enemies with native substrates in olive nurseries help to control phytonematodes?

Notes de la rédaction
Received 14 August 2018, accepted 19 November 2019, available online 17 december 2019
This article is distributed under the terms and conditions of the CC-BY License (http://creativecommons.org/licenses/by/4.0)
Résumé
La dispersion passive de champignons prédateurs par les substrats de culture peut-elle favoriser le biocontrôle des nématodes phytoparasites en pépinières d’oliviers ?
Description du sujet. Au Maroc, le développement d’une agriculture intensive dans le cadre du « Plan Maroc Vert » induira très probablement l’émergence de pathologies végétales, dont des nématoses. Dans le cas de l’olivier, la majorité des pépiniéristes utilisent des substrats constitués de sols d'origine indigène potentiellement contaminés en nématodes phytoparasites (NPP) et en champignons nématophages (CNP).
Objectifs. Le but de ce travail était de pouvoir évaluer la co-occurrence des NPP et des CNP dans les substrats ainsi que le potentiel antagoniste des CNP.
Méthode. La diversité des genres de NPP et de CNP a été caractérisée dans 305 échantillons de substrats prélevés dans 25 pépinières d'oliviers. L'activité fonctionnelle (nématicide et enzymatique) des CNP a été évaluée.
Résultats. Une grande diversité de NPP a été détectée, dont les 10 espèces les plus pathogènes à l’échelle mondiale. Les substrats se sont avérés riches en CNP qui diffèrent par leur activité prédatrice. L’analyse des communautés a révélé une partition nette entre NPP et CNP dans les substrats, confirmant l’antagonisme proie-prédateur. De plus, les patrons de NPP et de CNP dépendent de la stratégie trophique des CNP (saprophytes vs parasites obligatoires) et du mode d’antagonisme (prédation vs toxicité) impliqués. Mais les souches de CNP les plus performantes (croissance, sporulation, activité enzymatique) n'étaient pas forcément les plus efficaces contre les NPP. Par conséquent, la prédation et la compétition pourraient être un processus d'adaptation permettant au CNP de surmonter le stress environnemental et d'exploiter des sources nutritives alternatives ou supplémentaires.
Conclusions. L'utilisation des CNP indigènes offre des perspectives pour le développement de stratégies de gestion microbiologique des NPP respectueuses de l’environnement.
lutte biologique, communautés, écologie des sols, invasion, prédation
Abstract
Description of the subject. In Morocco, expanding agricultural sector under “Morocco Green Plan” will also introduce plant diseases and plant parasitic nematodes (PPN). Soil substrate used in most of the olive nurseries is potentially inhabited by PPN and natural enemies.
Objectives. This work assessed the co-occurrence of PPN and nematophagous fungi (NF) in substrates and potential of NF to suppress PPN.
Method. The diversity of PPN and NF genera was characterized in 305 substrate samples, isolated from 25 olive nurseries and functional activity (nematicidal and enzymatic) of NF was evaluated.
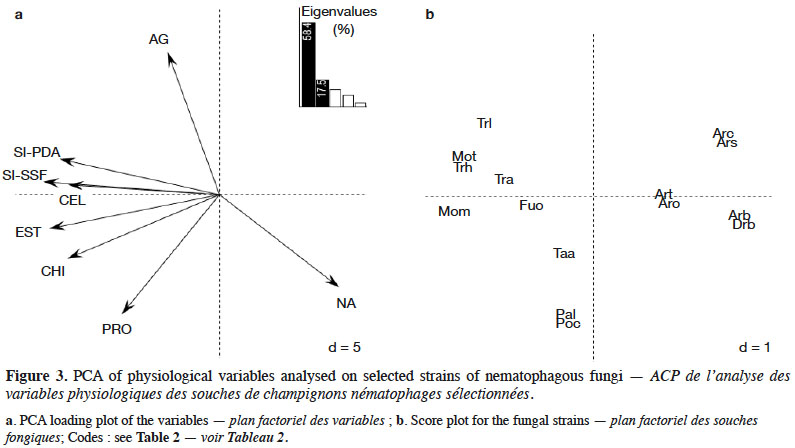
Results. High PPN diversity was detected in the isolated samples and substrates were differentiated by NF activity. Global partition between PPN and NF diversity patterns confirmed prey-predator antagonism. Moreover, PPN-NF patterns depend on the fungal survival (saprophytes vs obligate parasites) and antagonism (predation vs toxicity) processes. However, even the NF strains with maximum fitness (growth, sporulation, enzymatic activity) were not inevitably the most effective against PPN. Consequently, it was hypothesized that predation and competition could be an adaptation process of NF to overcome environmental stress and exploit alternative or supplementary nutrient sources.
Conclusions. The use of diverse native NF from cultivated and wild soils offer future prospects for the development of PPN management strategies in olive nurseries.
Table des matières
1. Introduction
1Soil borne fungi are one of the several microorganisms that act as antagonists to plant parasitic nematodes (PPN) (Poinar & Jansson, 1988). These fungi include endoparasitic fungi, predacious fungi, fungi parasitizing cysts and eggs of nematodes and fungi producing toxic metabolites against nematodes (Stirling, 1991). Nematophagous fungi (NF) utilize free-living nematodes and PPN as a nutrient and energy source. They are usually facultative saprobes but their saprophytic ability varies among different species. Initially, nematodes were considered the only nutrient source of predaceous fungi and hence were suggested for the biological control of PPN (Linford, 1937). Competition among various saprophytes in the soil rhizosphere to overcome nutrient deficiency should ultimately lead to enhanced nematode predation (Kerry, 2000).
2Predatory fungi can capture nematodes by elaborated traps, ranging from undifferentiated hyphae to complex constricting rings. Induction of saprophytic phase to predatory phase is triggered by several factors. Tridentaria implicans, Arthrobotrys anomala and A. botryospora capture nematodes by adhesion through sticky substance that covers them (Barron & Davidson, 1972; Barron, 1979). Nordbring-Hertz & Stålhammar-Carlemalm (1978) have deeply studied the nematodes predation by Arthrobotrys oligospora. Trapped nematodes struggle violently and fungus penetrates inside by perforating the cuticle. Then it develops an infection bulb that progressively invades the nematode, absorbing its contents and causing death within hours.
3Despite the fact that important research about soil microbiology had been conducted, interactions among different organisms in the soil remain little known. However, it is considered that fungi, bacteria, nematodes and other microorganisms compete for energy sources in the soil. Hyphomycete predators have the advantage to capture additional food source through their traps.
4Many NF species attack both parasitic and free-living nematode hosts and their densities in the rhizosphere are not related to the nematode populations. Previous studies about biological control of PPN assumed that activity of NF also increases with the rise in PPN populations (Linford & Oliveira, 1938; Duddington, 1960). However, it is difficult to confirm the level of NF dependency on nematodes for nutrition, especially with the species like Arthrobotrys oligospora that are successful soil saprophytes (Jaffee, 1992).
5Plant parasitic nematodes biocontrol is difficult in warm regions where nematode population densities are usually large due to host specificity and adaptation to environmental conditions. Generally, there are two limiting factors for biocontrol:
6– biocontrol through exotic agents rather than enhancing the efficiency of indigenous agents;
7– environmental impact on acclimatization of exotic agents.
8Consequently, prey-predator interactions and impact of the soil environment on exotic agents need to be evaluated, especially in the environments where native NF and PPN co-exist.
9Nematophagous fungi and PPN co-exist in nursery substrates used for breeding olive seedlings (Aït Hamza et al., 2017b; Aït Hamza et al., 2018). Aït Hamza et al. (2017b) demonstrated that NF in nursery substrates effectively killed Meloidogyne javanica juveniles and eggs.
10Considering that NF and PPN in olive nursery substrates are introduced with soil material from diverse habitats (open fields, forests, riverbanks), this study was designed to evaluate the co-occurrence of NF and PPN in substrates. Predation and lytic properties of NF were also analysed to find effective candidates for PPN biocontrol.
2. Materials and methods
2.1. Site description and sampling of olive root-cuttings
11Twenty-five commercial olive nurseries located in Jbala, Guerouane, Haouz and Souss regions of Morocco were selected to collect soil samples (Figure 1 & Table 1). Nurseries were selected based on cutting production, cultivar multiplication and substrates.
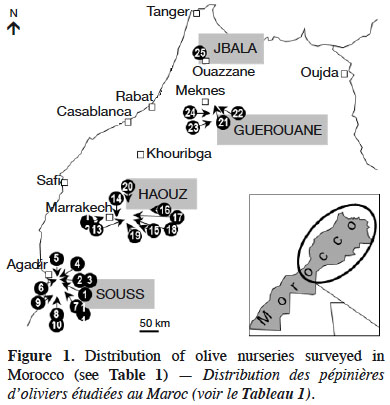
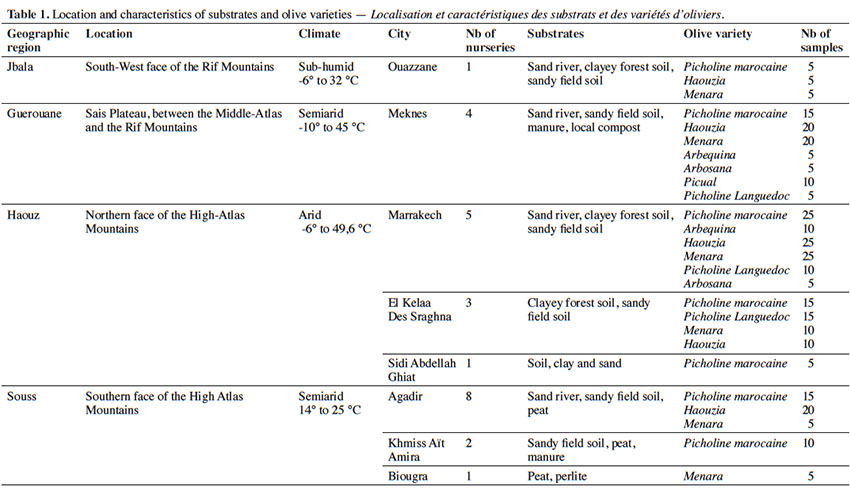
12Olive plants were grown in 2-to-3 litre plastic bags. Bags were filled with organic forest soils, sandy alluvial river bank soils, or loamy cropped soils (Table 1). Bags were further supplemented with fertilizer, sand, peat and animal manure. Plants were initially grown in plastic greenhouses and transplanted outside later on. Plants were watered with sprinkler and fertilized with Osmocote®, Everris Company™, at a dose of 45 g·plant-1.
13Five olive cuttings (Olea europaea subsp. europaea) were sampled for each grown variety from selected nurseries. Information about growing substrates and cultivars was recorded. In total, 305 olive root cuttings were collected and kept in a green house.
2.2. Nematode extraction and quantification
14To extract nematodes, substrate subsample of 250 cm3 was collected from the rhizosphere of each olive cutting and elutriation procedure (ISO 23611-4, 2007) was followed according to Oostenbrink (1960). Five cm3 aliquots were obtained from 25 ml stock suspensions to enumerate PPN of different orders including Aphelenchida (fungal feeders able to feed alternatively on plants), Dorylaimida, Triplonchida and Tylenchida under a stereomicroscope at 60X magnification. They were identified to the genus level based on dichotomous keys (Mai & Mullin, 1996) and their population levels were presented as per dm3 of fresh soil.
2.3. Isolation and characterization of nematophagous fungi
15Nematophagous fungi (NF) were isolated from RKN infested solid substrate samples by following soil sprinkling technique according to Santos et al. (1991). Media were directly powdered with soil to provide more concentrated base for fungal isolation than alternative Baermann technique. Moreover, sprinkling of cold soil (4 °C) on culture medium (37-40 °C) enhanced fungal isolation (Davet & Rouxel, 2000). Substrate aliquots of olive cuttings were air dried by spreading on a tray. Petri dishes containing water agar (WA 2% w/v) and antibiotics (0.05% chloramphenicol and 0.05% streptomycin sulphate) were sprinkled with 1 g of soil. Three replicates were prepared for each olive cutting. One milliliter suspension containing approximately 3,000 M. javanica second stage juveniles (J2) and 10 eggs was added to the Petri dishes (Aït Hamza et al., 2017a). Petri dishes were sealed with parafilm and incubated at room temperature (20-27 ºC) in darkness for two months. Petri dishes were examined daily during the first week of incubation and on weekly basis later on. Developed fungi were isolated under a microscope (60X magnification). Conidia or pieces of agar having NF mycelium or infected nematodes were transferred on Petri dishes containing Corn Meal Agar (CMA) or Potato Dextrose Agar (PDA) and antibiotics. After growing, fungi were transferred to a new culture medium plate for purification. Pure cultures were maintained on PDA or CMA plates at 4 °C in darkness for six months before regeneration.
16Slides were prepared from the pure NF culture and observed under dissecting microscope at 100X magnification. Specialized taxonomic keys were followed to assign genera (Philip, 2002; Yu et al., 2014), and species were confirmed by molecular characterization (Aït Hamza et al., 2017b).
2.4. Solid state fermentation and enzyme assays
17A substrate constituting sugarcane bagasse (50%), wheat bran (20%) and potato flour (10%) was used for solid-state fermentation (SSF) (Brand et al., 2000). Substrate was sterilized at 121 °C for 30 min and inoculated with 2x107 spores·g-1 after cooling. Initial pH and moisture contents were adjusted at 5.6 and 66%, respectively. Material (10 g) was packed into flasks and incubated at 25 °C.
18One-week old fermented material (5 g) was mixed with 50 ml of distilled water, homogenized for 1 min in Ultra-turax and pH was measured. Protease, chitinase, esterase and cellulase activities were determined according to Mukherjee et al. (2008) and reduced sugars were determined by spectrophotometry. International unit of cellulase activity is the amount of enzyme to release 1 μmol of glucose per min. Sample solution (2 ml) and DNS reagent (3 ml) were kept in a test tube to stir and then placed in a bath at 100 ºC for 5 min. The reaction was stopped in a frozen bath. A calibration curve of glucose was displayed from 0 to 1 g·l-1 and measured at 575 nm.
19Sporulation indices on PDA and SSF (SI-PDA and SI-SSF respectively), apical growth (AG) and nematicidal activity (NA) were evaluated according to Ait Hamza et al. (2017b).
2.5. Data analysis
20Co-Inertia Analysis (Dolédec & Chessel, 1994; Dray et al., 2003) was carried out for the abundance of PPN and NF. Statistical analyses were conducted in R 3.2.0 and multivariate analyses were performed by using R language (Dray & Dufour, 2007; R Development Core Team, 2011).
3. Results
3.1. Correspondance between plant-parasitic nematode and nematophagous fungal community patterns in olive nurseries of Morocco
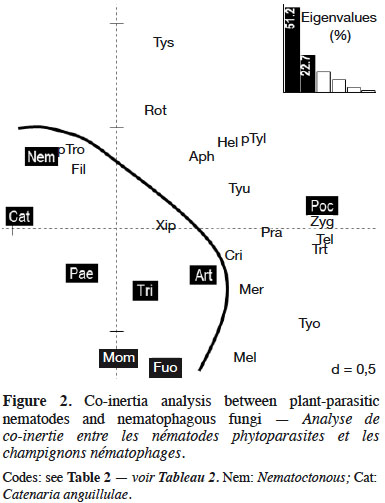
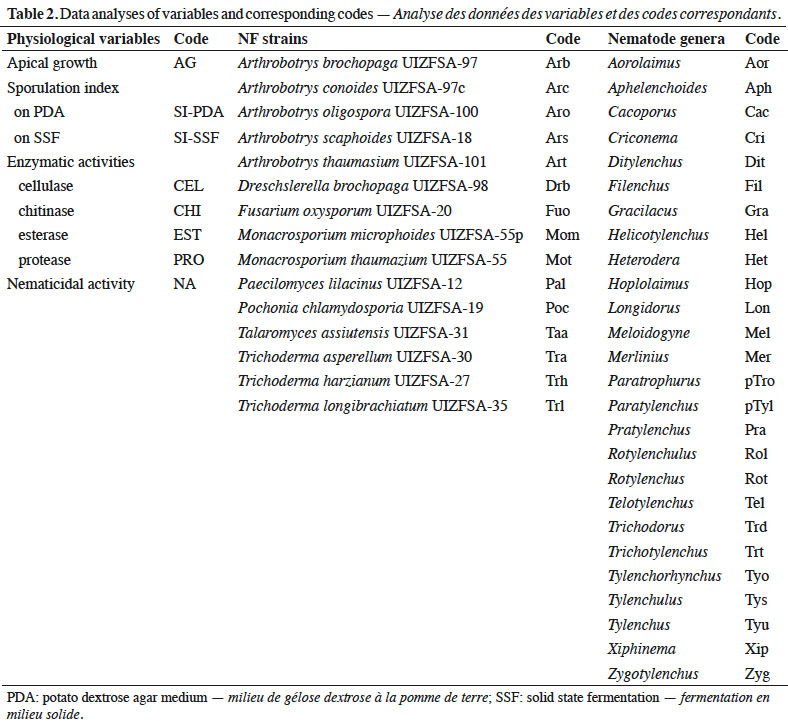
21Loading plot of the CIA analysis revealed a clear partition between NF (mostly in the left and bottom part of the plot) and PPN data (mostly in the right and above part of the plot) (Figure 2 and Table 2). Co-existence of NF Pochonia (Poc) and PPN Zygotylenchus (Zyg), Telotylenchus (Tel) and Trichotylenchus (Trt) is highlighted. Nematode genera Paratrophurus (pTro) and Filenchus (Fil) co-existed with NF genus Nematoctonous (Nem).
3.2. Hierarchy of functional traits
22Production of chitinase, esterase, protease, and cellulase enzymes has been very heterogeneous depending upon the strains (Figure 3 and Table 2). Overall, protease activity was the highest enzyme activity regardless of the strain, except M. microphoides and M. thaumasium, which exhibited significant cellulase activity.
23Unlike ANOVA, PCA analysis measures possible interactions between variables. Figure 3A indicated that cellulase (CEL), esterase (EST) and chitinase (CHI) activities, and sporulation indices (SI-PDA and SI-SSF), were correlated with PC1 axis (negative values). Apical growth (AG) and protease activity (PRO) were correlated with PC2 axis (positive and negative values respectively). Nematicidal activity (NA) was correlated with both PC1 and PC2 axes. Trichoderma (Tra, Trh and Trl) and Monacrosporium (Mom and Mot) strains were characterized through high sporulation indices and significant cellulase, esterase and chitinase activities. Pochonia chlamydosporia (Poc) and Paecilomyces lilacinus (Pal) strains exhibited significant protease activity, but low apical growth. Moreover, these strains, as well as Arthrobotrys (Arb, Arc, Aro, Ars and Art) and D. brochopaga (Drb) strains, were characterized by high nematicidal activity. Fusarium oxysporum (Fuo) and Talaromyces assiutensis (Taa) strains did not contribute to the PCA (Figure 3b).
4. Discussion
24Aim of the study was to examine co-occurrence of PPN and NF. Most of the information about fungal predation is based on agar plate experiments and limited data are available regarding predation in soils. Indeed, an ecological study on the co-occurrence of PPN and NF has never been conducted. Almost all rhizospheric microorganisms interact with each other. Strong antagonism between NF and PPN has been identified, except for Pochonia chlamydosporia that might have different niches. The clear partition between NF and PPN in olive nursery substrates can be explained by antagonistic effects among these faunas. Previous studies have also reported a multitude of fauna-fauna interactions in soil organisms (Giller, 1996). Considering that, nematodes and fungi are species of various trophic levels but interactions within and between fungi and nematodes are presumably the common point. In addition, indirect relationships between microorganisms are common characteristics of soil fauna interactions. Therefore, trophic guilds based concepts partly represent multitude fauna-fauna interactions in soils.
25Consequently, co-existence of NF and PPN might relate to the fact that each NF has a specific relation with PPN. For example, Catenaria anguillulae infects nematodes with its spores and spreads its vegetative hyphae inside infected nematodes. Nematoctonus leiosporus captures nematodes with adhesive spores and adhesive traps. Egg-parasitic fungi, such as Pochonia chlamydosporia and Paecilomyces spp., use appressoria to penetrate nematode eggs. Arthrobotrys spp. are more saprophytic than endoparasites and depend upon environmental conditions and presence of nematodes. Monacrosporium spp. regularly and spontaneously forms adhesive branches (Lopez-Llorca et al., 2006) whereas Trichoderma spp. and Fusarium oxysporum act with toxins (Sikora et al., 2008).
26There are two types of predation mechanisms:
27– a functional response that increases number of preys consumed by predators when prey density increases;
28– numerical response or change in the density of predators when prey numbers increase.
29Thompson (1924) reported that the killed prey is constant and independent of their density, while in contrast Nicholson & Baily (1935) have reported that the number of attacked preys depends on the parasite research area that is a constant and numbers of killed prey are directly proportional to their density. Errington (1946) demonstrated that vertebrates predation becomes important only when number of preys exceeds a safe level that is based on the maximum biotic niche. Moreover, opposition observed among some NF themselves can be explained according to Morin (1983) who indicated that predators could interpose in the interspecific competitions.
30Statistical co-existence of an egg-parasite (P. chlamydosporia) and different PPN species (Zygotylenchus spp., Pratylenchus spp. and Telotylenchus spp.) or co-existence of trapping fungus (Arhrobotrys spp.) with Criconema spp. and Merlinus spp. may relate to a delayed density dependant prey-predator interaction (increase in nematode numbers and a late increase of the fungus). Such co-existence between Pasteuria penetrans and root-knot nematodes has been demonstrated by Mateille et al. (2008). Previous studies also demonstrated that predatory fungi substantially reduced the populations of root-knot and citrus nematodes (Khan & Kim, 2005). On the other hand, Duddington (1960) proposed that three nematode genera might stimulate P. chlamydosporia production.
31Toxin-producing fungi, such as Trichoderma spp., continuously produce toxins regardless of the presence of nematodes. In that case, dynamics of PPN population do not respond to dependent prey-predator interactions with fungi. Antagonism between toxic fungi and nematodes is opportunistic and depends only on the presence of fungi and their adaptability to specific environment (e.g. forest soils and low temperatures). It also depends upon the specific susceptibility of nematodes to toxic compounds such as proteolytic enzymes (Sharon et al., 2001; Ahman et al., 2002).
32Resource competition in soils determines the structure of microbial communities (Lockwood & Filonow, 1981) as NF and PPN communities are affected by predation. Predation has a significant effect on the relative abundance of Arthrobotrys and Paecilomyces species but it does not cause any effect on the richness or diversity of PPN species. Taxa might be often under environmental constraints.
33As previously suggested (Brants et al., 2000; Sharon et al., 2001), protease and chitinase activities of filamentous fungi such as Trichoderma confirmed their potential as biocontrol agents against nematodes (Sharon et al., 2001). Proteolytic activity appears to correlate with nematode controlling capabilities (Ahman et al., 2002) and Trichoderma is considered to use subtilisin against nematodes (Sharon et al., 2001). Trypsin-like proteases have also been identified for their nematotoxic activities (Sharon et al., 2001; Ahman et al., 2002). Our results depicted that enzyme activities vary among species and NF species exhibit significant enzyme activities. These activities offer a promising basis for developing alternative nematode control methods. During the study, most of the strains exhibited activities of four targeted enzymes. It suggests that NF can affect embryo development of nematodes by degrading their chitino-protein layer through enzymatic tools. This is consistent with the hypothesis that main nematicidal activity of Trichoderma spp. occurs in soil rather than in roots (Brants et al., 2000). Trichoderma, Fusarium and Monacrosporium species with key enzyme activities (esterase, cellulase) justify their lethal effects on nematodes. In the terrestrial environment features like production of enzymes, bioactive substances and rapid development make Trichoderma spp. a potential agent in the agro-food sector and a material of choice for industrial exploitation (Prieto et al., 1997).
34On the other hand, Arthrobotrys and Dreschslerella brochopaga seem less efficient in terms of spore and enzyme production but they can efficiently trap nematodes through specific mycelial structures. Pochonia chlamydosporia and Paecilomyces lilacinus are more active on nematode eggs via protease secretion as reported by Yang et al. (2013). The case of Talaromyces assiutensis remains elusive so far as its poor physiological capacities analysed in this study do not correspond to its high nematicidal efficiency as previously observed by Aït Hamza et al. (2017).
5. Conclusions
35During this study, significant antagonist processes were observed between co-existing NF-PPN species in olive nursery substrates. However, it was clearly established that PPN-NF interactions depend on antagonism processes (predation vs toxicity), different species and environmental conditions. Future studies should focus on exploring NF capabilities to control specific PPN and co-existence of NF communities to develop multi-NF control strategies under different environmental conditions.
Acknowledgements
36Institut de Recherche pour le Développement (Marseille, France) supported this research under the Ph.D grant. It was co-funded by PESTOLIVE project: Contribution of olive history for the management of soil-borne parasites in the Mediterranean Basin (ARIMNet action KBBE 219262), and BIONEMAR project: Development of fungal bionematicides for organic production in Morocco (PHC-Toubkal action 054/SVS/13).
Bibliographie
Ahman J. et al., 2002. Improving the pathogenicity of a nematode-trapping fungus by genetic engineering of a subtilisin with nematotoxic activity. Appl. Environ. Microbiol., 68, 3408-3415, doi.org/10.1128/aem.68.7.3408-3415.2002
Aït Hamza M. et al., 2017a. Diversity of root-knot nematodes in Moroccan olive nurseries and orchards: does Meloidogyne javanica disperse according to invasion processes? BMC Ecol., 17(1), 41, doi.org/10.1186/s12898-017-0153-9
Aït Hamza M. et al., 2017b. Diversity of nematophagous fungi in Moroccan olive nurseries: highlighting prey-predator interactions and efficient strains against root-knot nematodes. Biol. Control, 114, 14-23, doi.org/10.1016/j.biocontrol.2017.07.011
Aït Hamza M. et al., 2018. Diversity of plant-parasitic nematode communities associated with olive nurseries in Morocco: origin and environmental impacts. Appl. Soil. Ecol., 124, 7-16, doi.org/10.1016/j.apsoil.2017.10.019
Barron G., 1979. Nematophagous fungi: a new Arthrobotrys with nonseptate conidia. Can. J. Bot., 57, 1371-1373, doi.org/10.1139/b79-170
Barron G. & Davidson J., 1972. Nematophagous Hyphomycetes: Arthrobotrys anomala sp. nov. Can. J. Bot., 50, 1773-1774, doi.org/10.1139/b72-220
Brand D., Pandey A., Roussos S. & Soccol C.R., 2000. Biological detoxification of coffee husk by filamentous fungi using a solid state fermentation system. Enzyme Microb. Technol., 27, 127-133, doi.org/10.1016/s0141-0229(00)00186-1
Brants A., Brown C.R. & Earle E.D., 2000. Trichoderma harzianum endochitinase does not provide resistance to Meloidogyne hapla in transgenic tobacco. J. Nematol., 32(3), 289.
Davet P. & Rouxel F., 2000. Detection and isolation of soil fungi. Enfield, NH, USA: Science Publisher Inc.
Dolédec S. & Chessel D., 1994. Co-inertia analysis: an alternative method for studying species-environment relationships. Freshwater Biol., 31, 277-294, doi.org/10.1111/j.1365-2427.1994.tb01741.x
Dray S., Chessel D. & Thioulouse J., 2003. Co-inertia analysis and the linking of ecological tables. Ecology, 84(11), 3078-3089, doi.org/10.1890/03-0178
Dray S. & Dufour A.B., 2007. The ade4 package: implementing the duality diagram for ecologists. J. Stat. Software, 22, 1-20, doi.org/10.18637/jss.v022.i04
Duddington C., 1960. Biological control - predaceous fungi. In: Sasser J.N. & Jenkins W.R., eds. Nematology, fundamentals and recent advances with emphasis on plant parasitic and soil forms. Chapel Hill, NC, USA: University of North Carolina Press, 461-465.
Errington P.L., 1946. Predation and vertebrate populations. Q. Rev. Biol., 21, 144-177.
Giller P.S., 1996. The diversity of soil communities, the ‘poor man's tropical rainforest’. Biodivers. Conserv., 5, 135-168, doi.org/10.1007/bf00055827
ISO 23611-4, 2007. Soil quality - Sampling of soil invertebrates - Part 4: Sampling, extraction and identification of soil-inhabiting nematodes. Geneva, Suisse: ISO.
Jaffee B., 1992. Population biology and biological control of nematodes. Can. J. Microbiol., 38, 359-364, doi.org/10.1139/m92-061
Kerry B.R., 2000. Rhizosphere interactions and the exploitation of microbial agents for the biological control of plant-parasitic nematodes. Annu. Rev. Phytopathol., 38, 423-441, doi.org/10.1146/annurev.phyto.38.1.423
Khan Z. & Kim Y., 2005. The predatory nematode, Mononchoides fortidens (Nematoda: Diplogasterida), suppresses the root-knot nematode, Meloidogyne arenaria, in potted field soil. Biol. Control, 35, 78-82, doi.org/10.1016/j.biocontrol.2005.05.015
Linford M.B., 1937. Stimulated activity of natural enemies of nematodes. Science, 85, 123-124, doi.org/10.1126/science.85.2196.123
Linford M. & Oliveira J., 1938. Potential agents of biological control of plant-parasitic nematodes. Phytopathology, 28, 14.
Lockwood J.L. & Filonow A.B., 1981. Responses of fungi to nutrient-limiting conditions and to inhibitory substances in natural habitats. In: Alexander M., ed. Advances in microbial ecology. Vol. 5. Springer, doi.org/10.1007/978-1-4615-8306-6_1
Lopez-Llorca L.V., Jansson H.B., Vicente J.G.M. & Salinas J., 2006. Nematophagous fungi as root endophytes. In: Schulz B., Boyle C. & Sieber T.N., eds. Microbial root endophytes. Berlin/Heidelberg, Germany: Springer, 191-206, doi.org/10.1007/3-540-33526-9_11
Mai W.F. & Mullin P.G., 1996. Plant-parasitic nematodes: a pictorial key to genera. 5th ed. New York, NY, USA: Comstock Publishing Associates.
Mateille T., Cadet P. & Fargette M., 2008. Control and management of plant parasitic nematode communities in a soil conservation approach. In: Ciancio A. & Mukerji K.G., eds. Integrated management and biocontrol of vegetable and grain crops nematodes. Springer, 79-97, doi.org/10.1007/978-1-4020-6063-2_4
Morin P.J., 1983. Predation, competition, and the composition of larval anuran guilds. Ecol Monogr., 53, 119-138, doi.org/10.2307/1942491
Mukherjee A.K., Adhikari H. & Rai S.K., 2008. Production of alkaline protease by a thermophilic Bacillus subtilis under solid-state fermentation (SSF) condition using Imperata cylindrical grass and potato peel as low-cost medium: characterization and application of enzyme in detergent formulation. Biochem. Eng. J., 39, 353-361, doi.org/10.1016/j.bej.2007.09.017
Nicholson A.J. & Bailey V.A., 1935. The balance of animal populations. Part I. Proc. Zool. Soc. London, 105, 551-598, doi.org/10.1111/j.1096-3642.1935.tb01680.x
Nordbring-Hertz B. & Stålhammar-Carlemalm M., 1978. Capture of nematodes by Arthrobotrys oligospora, an electron microscope study. Can. J. Bot., 56, 1297-1307, doi.org/10.1139/b78-146
Oostenbrink M., 1960. Estimating nematode populations by some selected methods. In: Sasser J.N. & Jenkins W.R., eds. Nematology. Chapel Hill, NC, USA: The University of North Carolina Press, 85-102.
Poinar G.O. Jr. & Jansson H.B., eds, 1988. Diseases of nematodes, vol. I and II. Boca Raton, FL, USA: CRC Press.
Prieto A. et al., 1997. Structure of complex cell wall polysaccharides isolated from Trichoderma and Hypocrea species. Carbohydr Res., 304(3-4), 281-291, doi.org/10.1016/s0008-6215(97)00239-5
R Development Core Team, 2011. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Santos M., Ferraz S. & Muchovej J., 1991. Detection and ecology of nematophagous fungi from Brazil soils. Nematol. Bras., 15, 121-134.
Sharon E. et al., 2001. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Phytopathology, 91(7), 687-693, doi.org/10.1094/phyto.2001.91.7.687
Sikora R.A. et al., 2008. Mutualistic endophytic fungi and in-planta suppressiveness to plant parasitic nematodes. Biol. Control, 46(1), 15-23, doi.org/10.1016/j.biocontrol.2008.02.011
Stirling G., 1991. Conservation and enhancement of naturally occurring antagonists and the role of organic matter. In: Biological control of plant parasitic nematodes. Progress, Problems and Prospects. Wallingford, UK: CAB International.
Thompson W., 1924. La théorie mathématique de l’action des parasites entomophages et le facteur du hasard. Ann. Fac. Sci. Marseille, 2, 69-89.
Yang J., Liang L., Li J. & Zhang K.-Q., 2013. Nematicidal enzymes from microorganisms and their applications. Appl. Microbiol. Biotechnol., 97, 7081-7095, doi.org/10.1007/s00253-013-5045-0
Yu Z., Mo M., Zhang Y. & Zhang K.Q., 2014. Taxonomy of nematode-trapping fungi from Orbiliaceae, Ascomycota. In: Zhang K.Q. & Hyde K.D., eds. Nematode-trapping fungi. Dordrecht, The Netherlands: Springer, 41-210, doi.org/10.1007/978-94-017-8730-7_3
