


- Portada
- Volume 21 (2017)
- numéro 3 : Atelier "Fonctions agro-écosystémiques ...
- Renouvellement du carbone profond des sols cultivés : une estimation par compilation de données isotopiques


Vista(s): 2012 (33 ULiège)
Descargar(s): 457 (11 ULiège)

Renouvellement du carbone profond des sols cultivés : une estimation par compilation de données isotopiques

Notes de la rédaction
Reçu le 22 mars 2016, accepté le 6 septembre 2016
Cet article est distribué suivant les termes et les conditions de la licence CC-BY (http://creativecommons.org/licenses/by/4.0/deed.fr)
Résumé
Description du sujet. L'article présente une estimation du renouvellement à moyen terme (dizaines d'années) du carbone profond des sols cultivés.
Objectifs. Le stockage de carbone sous forme organique dans les sols est un moyen de limiter l'augmentation du CO2 global et le réchauffement climatique. L'objectif est de proposer une première quantification du renouvellement des horizons profonds (> 30 cm) qui contiennent environ la moitié du carbone organique des sols.
Méthode. Nous avons effectué une synthèse d'études de la dynamique du carbone de 41 profils où le renouvellement du carbone est quantifié grâce au traçage isotopique par les abondances naturelles en carbone-13.
Résultats. Le renouvellement du carbone profond est en moyenne quatre fois plus lent qu'en surface, mais le flux de renouvellement n'est pas négligeable. La distribution moyenne du flux annuel d'apport de carbone aux matières organiques est de 81 % dans l'horizon 0 - 30 cm et 19 % dans la couche 30 - 100 cm, avec une erreur standard de ± 4 %. À plus long terme (20 ans), le sous-sol (30 - 100 cm) contient en moyenne 23 % du carbone récent du sol.
Conclusions. Les matières organiques profondes ne doivent pas être négligées dans les bilans de C et N en réponse à différentes pratiques agricoles, et nous proposons une approximation simple pour ces bilans. Dans le futur, les estimations des évolutions temporelles du stock de carbone profond des sols et de leurs facteurs de variation devront s'appuyer sur les essais agronomiques de longue durée ou la modélisation calée sur des sites fortement instrumentés, et être accompagnées de descriptions pédologiques détaillées.
Abstract
Turnover of deep organic carbon in cultivated soils: an estimate from a review of isotope data
Description of the subject. This study is an estimate of medium-term renewal (decades) of deep carbon in cultivated soils.
Objectives. Sequestration of organic carbon in soils is a way to mitigate the global rise in CO2 and warming. Deep horizons (> 30 cm) contain about half of the soil organic carbon but, in agriculture, deep organic carbon and nitrogen are often neglected when the balance of these elements is calculated. Our goal in this study was to propose a quantification of deep C turnover.
Method. We collected data from 41 profiles, where C turnover was studied using the natural 13C labeling technique. The studied cropping systems were basically tropical and temperate C4 plant monocultures.
Results. Deep carbon (30 – 100 cm) turnover was on average four times slower than in topsoil (0 – 30 cm), but renewal was significant. The average depth-distribution of the carbon flux to organic matter build-up was 81% in the layer at the 0 – 30 cm level and 19% in the layer at 30 – 100 cm, with a standard error of ± 4%. Over the longer term (20 years), subsoil at 30 – 100 cm contained on average 23% of the recent soil carbon accumulated in the first meter.
Conclusions. Deep organic matter should not be overlooked when considering the balance of C and N and we therefore propose a simple method for an initial first assessment. In the future, further estimates of the temporal evolution of the deep carbon stock and its factors of variation should be based on long-term agronomic experiments or modeling parameterized on heavily instrumented sites, and should be accompanied by detailed pedological descriptions.
Tabla de contenidos
1. Introduction
1La question de la dynamique du carbone organique profond des sols est posée actuellement par la communauté scientifique qui évalue le cycle planétaire du carbone en lien avec l'évolution du climat. La taille du réservoir de carbone des sols de la planète est évaluée entre 1 500 Gt C et 2 400 Gt C selon les auteurs et les profondeurs prises en compte (Hiederer et al., 2011 ; Ciais et al., 2013), hors zone de permafrost (Koven et al., 2011), ce qui en fait un puits ou une source potentielle énorme par rapport à l'augmentation du CO2 atmosphérique (+ 4,4 Gt C en 2015). Or, à l'échelle planétaire comme à l'échelle de la France, la moitié du carbone est située en dessous de 30 cm de la surface du sol (Jobbagy et al., 2000 ; Hiederer et al., 2011 ; Mulder et al., 2016). Même si les teneurs en carbone sont plus basses qu'en surface, la masse volumique apparente plus élevée et l'épaisseur parfois importante du sol font que les stocks 0 - 30 et 30 - 100 cm sont du même ordre de grandeur.
2L'incertitude sur la dynamique du carbone des sols est souvent invoquée pour expliquer la relative mauvaise représentation du carbone des écosystèmes dans les modèles globaux, qu'il s'agisse de représenter les flux nets, le turnover ou le stock (Ahlström et al., 2012 ; Carvalhais et al., 2014). En effet, la dynamique du carbone superficiel est relativement bien quantifiée, en particulier grâce aux expérimentations de longue durée, dont les plus anciennes ont été initiées à la fin du 19e siècle, alors qu'il reste des interrogations majeures sur l'estimation du renouvellement du carbone des sols situé en dessous des 30 premiers centimètres et les processus en jeu (Harper et al., 2013), ce qui en fait une source d’incertitude majeure pour les prévisions sur l’évolution du cycle global du carbone et du climat.
3Plusieurs auteurs ont démontré que les mécanismes présidant à la préservation ou à la biodégradation peuvent s'exprimer avec des intensités différentes en surface et en profondeur (Fontaine et al., 2007 ; Salomé et al., 2010). En agriculture, la partie profonde des sols est particulièrement surveillée et gérée pour ses propriétés physiques, mais en général peu considérée en tant que réserve de matière organique, ou comme source ou puits d'éléments nutritifs. Les modèles opérationnels du bilan organique utilisés en agriculture, comme RothC (Coleman et al., 1995), ou AMG (Andriulo et al., 1999 ; Bouthier et al., 2014) ne considèrent d'ailleurs que la couche superficielle (0 - 30 cm dans l'Union européenne), alors que la distribution du carbone en fonction de la profondeur est un continuum, à la seule exception de l'horizon Ap où la distribution du C peut être homogène lorsqu'il y a, ou qu'il y a eu dans le passé, labour par retournement.
4Le renouvellement du carbone profond est lent, comme en atteste la datation au 14C : l'âge apparent 14C, proche de l'âge moyen du carbone, atteint couramment 2 500 ans à 1 m de profondeur (Mathieu et al., 2015). Ceci rend très difficile l'étude de sa dynamique par le suivi temporel des stocks. Les flux bruts sont également difficilement mesurables. Concernant les flux entrants, il s'agit essentiellement de mortalité racinaire, de rhizodéposition et de flux verticaux de matière organique soluble, qui sont très difficiles à estimer : le renouvellement racinaire peut être quantifié en longueur ou volume de racines, mais plus difficilement en masse et le caractère transitoire des rhizodépôts et exsudats les rend insaisissables sans traçage isotopique (Balesdent et al., 2011).
5S'agissant du flux sortant du réservoir organique, principalement la minéralisation sous forme de CO2, sa quantification in situ est rendue très délicate voire impossible car l'efflux de CO2 comprend également la respiration autotrophe racinaire, qui est neutre pour le bilan organique du sol, mais souvent supérieure à la respiration hétérotrophe des décomposeurs de matière organique (Hanson et al., 2000). Enfin, il est quasiment impossible d’accéder à la minéralisation du C profond sans induire une perturbation majeure des processus par la mise en place de capteurs, l'exclusion des racines ou le prélèvement de sol dans les horizons profonds.
6Une méthode de choix pour l'investigation de la dynamique du carbone est le traçage isotopique naturel : les variations d'abondance naturelle en 13C, un isotope stable du carbone, peuvent être exploitées dans le cas de cultures de plantes à cycle photosynthétique C4 établies sur des sols ayant porté auparavant des couverts de végétation C3 (Cerri et al., 1985). Les plantes C4 les plus couramment cultivées sont le maïs, le sorgho, le mil, la canne à sucre et des plantes à vocation énergétiques comme Miscanthus et Panicum virgatum (switchgrass). La différence de rapport 13C/12C entre plantes C3 et C4 est importante, si bien que, après t années de changement de végétation, la proportion de carbone organique provenant de la nouvelle végétation peut être déterminée à partir du rapport 13C/12C organique du sol. L'évolution de ce rapport en fonction du temps t est une mesure directe du renouvellement du carbone du sol.
7L'objectif de cette étude est de quantifier la répartition verticale des flux entrants, la variation des vitesses de renouvellement en fonction de la profondeur et, finalement, la contribution du carbone qui se trouve en profondeur, au pool de carbone « actif » des sols. Nous avons effectué, pour ce faire, la méta-analyse des données de 41 sites extraites de 25 études qui ont estimé le renouvellement du carbone le long de profils de sols cultivés au moyen du traçage isotopique par les abondances naturelles en 13C.
2. Matériel et méthodes
2.1. Sites et études sélectionnées
8La technique du traçage par les abondances naturelles en carbone 13C fait appel à deux parcelles qui ont porté la même végétation avant la mise en place des monocultures : la parcelle sous monoculture C4 et une parcelle de référence maintenue sous la végétation initiale ou mise en même temps sous végétation C3.
9Les études ont été sélectionnées selon plusieurs critères : la durée de la monoculture doit être connue précisément, le sol doit avoir été échantillonné jusqu'à au moins 70 cm ou à la roche mère, les concentrations en carbone de la parcelle C4 doivent avoir été mesurées. Nous avons compilé les données publiées de 41 situations, tirées de 25 références (Annexe 1), où a varié la durée des monocultures de plantes C4 établies sur sols ayant porté au préalable des végétations de type C3. La base de données comporte quatre grands types de systèmes de culture (Tableau 1). Les principaux grands types de sols (IUSS Working Group WRB, 2014) représentés sont : cambisols (non andic inceptisols) (n = 9), ferralsols et acrisols (oxisols et ultisols) (n= 9), andosols (n = 8), luvisols (alfisols) (n = 7), chernozems (mollisols) (n = 3), gleysols (n = 3), podzols (n = 2).
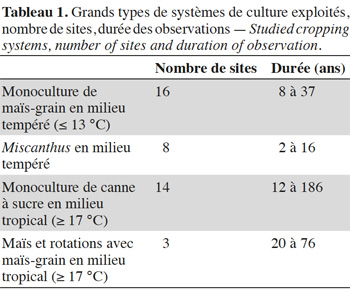
2.2. Calculs
10Le traçage isotopique permet, pour tout échantillon, de calculer la proportion f de carbone provenant de la plante cultivée depuis le début de la monoculture, c'est-à-dire de carbone plus jeune que t années. Le reste (1-f) est la proportion de carbone provenant des matières organiques présentes dans le sol avant le changement de végétation, qui n'a pas été minéralisée. Les stocks de carbone total, la proportion de carbone nouveau, provenant de la culture et les stocks de carbone nouveau ont été calculés de la manière suivante. Pour chaque couche (z1, z2), C est le stock de carbone de la couche (kg C·m-2), f est la proportion de carbone nouveau provenant de la culture et Cnouv est le stock de carbone nouveau (kg C·m-2). Les valeurs de C, f et Cnouv sont soit fournies par les auteurs, soit calculées à partir des concentrations en carbone [C] (g C·kg-1) et de la masse volumique apparente , comme suit :

11où est fournie par les auteurs, ou calculée (Alexander, 1980). Les éléments grossiers sont négligés. Les variables f et Cnouv sont calculées d'après l'équation de Balesdent et al. (1996) qui relie la composition isotopique du carbone organique à la proportion de carbone provenant de la nouvelle végétation de type C4 :
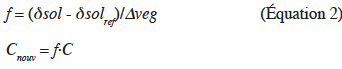
12où δsol et δsolrefsont les rapports 13C/12C du carbone de la couche considérée dans le sol étudié et le sol de référence, respectivement, et ∆veg est la différence des rapports 13C/12C entre la végétation C4 et la végétation C3 de référence. En raison des enrichissements naturels en 13C avec la profondeur, la valeur de δsolref est obtenue pour chaque couche à la même profondeur. L'équation 2 prend en compte les sources d'enrichissement isotopique autres que le changement de végétation, soit les variations isotopiques des végétations du passé et celles résultant de la biodégradation, et donne une estimation sans biais de f si ces deux effets sont identiques dans les deux profils (Balesdent et al., 1996).
13Afin de comparer les sites entre eux, nous avons intégré selon z les stocks cumulés C(0, z), Cnouv(0, z), puis calculé C et f par intervalle de 10 cm jusqu'à 100 cm par simple interpolation linéaire des stocks cumulés. Dans quelques cas de couches manquantes, les concentrations en carbone ont été interpolées à partir des couches supérieure et inférieure. Dans le cas de sites décrits jusqu'à une profondeur inférieure à 100 cm (le plus souvent 80 cm), nous avons procédé à une extrapolation pour rapporter le stock de carbone des couches observées au stock de la profondeur de référence 0 - 100 cm : nous avons extrapolé C(0, 100 cm) à partir de la profondeur maximale décrite zmax selon la régression C(0, 100 cm) = a * C(0, zmax) + b paramétrée pour l'ensemble des sites.
2.3. Analyses statistiques
14Les régressions sont effectuées en utilisant le module d'ajustement par modèle linéaire lm du logiciel R.
3. Résultats
3.1. Vitesses de renouvellement et âge du carbone en surface et en profondeur.
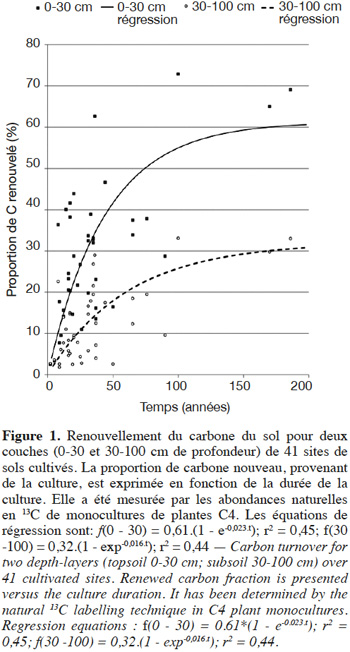
15La figure 1 présente la proportion de carbone nouveau f en fonction du temps t, c'est-à-dire le renouvellement du carbone, dans les deux couches 0 - 30 et 30 - 100 cm. Elle montre pour la couche supérieure quelques grandes tendances connues de la dynamique du carbone des sols. Le renouvellement du carbone est plus rapide dans les premières années, en raison de la présence de fractions rapidement décomposables et renouvelables, puis est de plus en plus lent, sans atteindre 100 % même après plusieurs siècles, en raison de la présence de carbone résistant ou protégé de la minéralisation à très long terme. L'âge médian du carbone (temps correspondant au renouvellement de 50 %) est de l'ordre de 100 ans pour la couche 0 - 30 cm. Le renouvellement du carbone est bien sûr très variable entre les sites. La variabilité, non discutée ici, est d'abord liée à trois facteurs : les phases minérales présentes, avec les minéraux comme les argiles ou les allophanes ralentissant le renouvellement ; le climat, avec un renouvellement plus rapide en conditions tropicales ; la différence de dynamique du carbone entre la monoculture étudiée et le système ou l'usage qui la précédait, qu'il s'agisse du changement des flux de restitution de carbone au sol ou des vitesses de minéralisation. Le renouvellement en profondeur est beaucoup plus lent. Sur chaque site, la proportion de carbone nouveau décroît progressivement avec la profondeur, jusqu'à une valeur de quelques % à 100 cm. Nous avons exprimé le renouvellement de la couche 30 - 100 cm des mêmes sites (Figure 1). En moyenne, la proportion de carbone nouveau dans la couche 30 - 100 cm est trois fois plus faible (2,9 avec un écart-type de 1,9) que dans la couche 0 - 30 cm. Des régressions exponentielles de type f = a*(1-e-k.t) ont été effectuées sur l'ensemble des points (Figure 1). Elles peuvent être interprétées comme décrivant grossièrement le renouvellement de la manière suivante : une proportion (a) du carbone est sujette à renouvellement, le reste (1 - a) étant stable à cette échelle de temps (200 ans) ; le taux moyen de renouvellement annuel de la partie renouvelée est k (an-1) ; il s'agit de renouvellement par les nouveaux apports par la végétation qui compensent des départs par minéralisation et migration. À partir de ces régressions, on estime que le temps nécessaire pour renouveler 20 % du carbone de l'horizon est en moyenne de l'ordre de trois à quatre fois plus long (environ 60 ans) pour 30 - 100 cm que pour 0 - 30 cm (environ 17 ans). Plusieurs hypothèses ont été avancées pour expliquer le renouvellement plus lent en profondeur (Rumpel et al., 2011 ; Guenet et al., 2013). Le carbone stable à l'échelle du millénaire est en proportion relative plus importante en profondeur qu'en surface et, à la durée de vie du carbone in situ, il faut ajouter le temps nécessaire au transport de carbone depuis la surface vers la profondeur, par pédoturbation, bioturbation ou lixiviation (Elzein et al., 1995 ; Guenet et al., 2013). Enfin, la biodégradation peut être plus lente qu'en surface, en raison de plus fortes interactions organo-minérales (Salomé et al., 2010) ou de moindre capacité des micro-organismes à dégrader les matières organiques car ces derniers reçoivent moins de ressources énergétiques de substrats frais (Fontaine et al., 2007). Ces hypothèses doivent être considérées comme en interaction entre elles, aucune n'excluant les autres.
3.2. Distribution verticale du carbone organique total
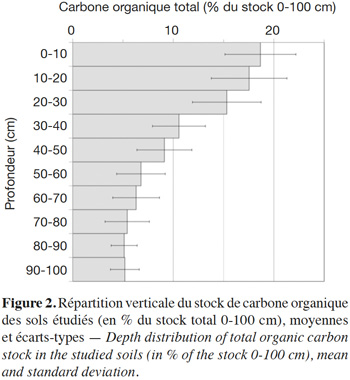
16La mesure des concentrations en carbone et de la densité apparente permet de calculer la répartition verticale du stock de carbone (Figure 2). Les concentrations volumiques en carbone sont en moyenne trois fois plus faibles à 1 m que dans l'horizon A. Les concentrations sont très semblables au sein de la couche 0-30 cm car la plupart des sites ont été labourés soit régulièrement, soit occasionnellement, ce qui homogénéise les teneurs en carbone jusqu'à la profondeur de labour (20 à 30, parfois 40 cm). Les horizons profonds (30-100 cm) contiennent en moyenne 48 % du carbone organique total du premier mètre, chiffre très proche des estimations à l'échelle de la France métropolitaine (Mulder et al., 2016) ou de la planète (Jobbagy et al., 2000). Au sein de l'ensemble des sols cultivés étudiés, cette proportion est assez variable (1σ = 10 %) et il est attendu que l'accumulation de carbone dans les horizons les plus profonds soit plus dépendante du type pédologique que de la culture elle-même. En effet, les travaux de Mathieu et al. (2015) ont montré au niveau de la planète que le degré de stabilisation (l'âge) du carbone profond est plus dépendant du type de sol que du climat ou de l'usage des terres.
3.3. Distribution verticale du carbone provenant de la culture
17Nous avons estimé pour chaque couche le stock de carbone récent, à partir des abondances en 13C et de la concentration volumique en carbone, et calculé pour chaque profil la répartition verticale de ce carbone récent, en % du stock de carbone récent de l'ensemble du profil (méthode en Annexe 1). Ces données sont regroupées par culture sur la figure 3. Le rapport du carbone nouveau dans la couche 30-100 cm sur le carbone nouveau de l'ensemble 0-100 cm est en moyenne de 0,24 pour le maïs, 0,22 pour Miscanthus et 0,36 pour la canne à sucre.
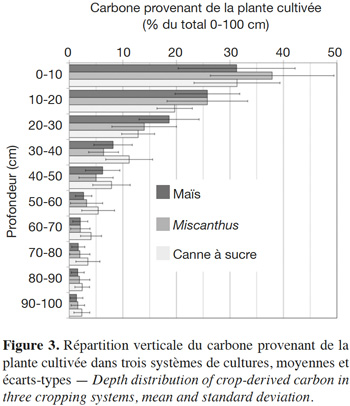
18La proportion du carbone nouveau localisé en profondeur dépend de plusieurs variables, notamment le temps d'observation et l'usage des sols. Ainsi, elle est plus importante pour les temps longs et dans les systèmes sous canne à sucre que sous cultures de maïs ou de Miscanthus (p < 0,001), ce qui est visible sur la figure 3. La variation peut être expliquée par d'autres facteurs que la distribution des restitutions de carbone ou les enracinements. En effet, le jeu de données présente une covariance statistique entre la nature de la culture et le climat bien sûr, mais aussi entre la nature et la durée de la monoculture. La durée médiane des études est de 11, 26 et 47 ans pour Miscanthus, maïs et canne à sucre, respectivement. Plusieurs facteurs peuvent ainsi expliquer l'accumulation supérieure que nous observons en profondeur sous canne à sucre. La durée d'observation étant plus importante, l'accumulation relative en profondeur est supérieure. D'autre part, le brûlage des feuilles de canne a pu être largement pratiqué, notamment dans le passé, réduisant d'autant les apports en surface.
19Pour analyser plus finement la dépendance du temps dans les systèmes sous Miscanthus et maïs, nous avons choisi comme indicateur unique d’incorporation le rapport R du stock de carbone nouveau de la couche profonde sur le stock de carbone nouveau de l'ensemble du profil :
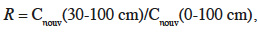
20et nous avons analysé sa variation par un modèle de régression linéaire et une analyse de variance. L'équation de régression linéaire en fonction du temps et du système de culture est la suivante :
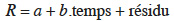
21avec a, dépendant du type de culture, b commun.
22L'estimation des paramètres est donnée au tableau 2. Les trois systèmes comparés sont très semblables. In fine, la régression linéaire simple, unique pour les systèmes en maïs et Miscanthus, en donne une bonne description :

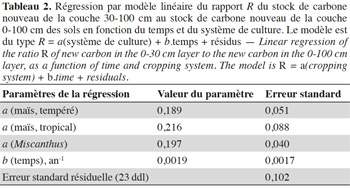
23L'ordonnée à l'origine (0,19) est assortie d'une erreur standard de 0,035 et la pente (0,0020 an-1) d'une erreur standard de 0,0013 an-1. L'erreur standard résiduelle est de 0,10, avec 25 degrés de liberté. À long terme (20 ans), la proportion du carbone nouveau est répartie en 77 % ± 4 % en surface et 23 % ± 4 % (équation 3). En d'autres termes, la quantité de carbone d'âge inférieur à 20 ans de la couche 30-100 cm est égale à 0,30 ± 0,07 fois celle de la couche 0-30 cm.
3.4. Distribution verticale des apports de carbone aux matières organiques du sol
24On peut, à partir de cet ensemble de sites, faire des estimations de la répartition verticale des flux d'apports de carbone aux matières organiques. Ceci ne peut être fait individuellement sur chaque site, car chacun a une durée d'observation différente de celle des autres, mais cela peut être fait pour l'ensemble, à partir de la cinétique moyenne d'évolution du rapport entre le carbone nouveau dans chaque couche et le carbone nouveau dans l'ensemble du profil. En effet, la répartition verticale du carbone nouveau est dépendante du temps dans le jeu de données de l'ensemble des sites (Équation 3). Ceci peut être dû à deux phénomènes indépendants :
25– le transport progressif de carbone vers la profondeur par bioturbation, lixiviation ou lessivage,
26– la biodégradation in situ du C nouveau, plus lente en profondeur qu'en surface.
27La dispersion dans le jeu de données ne permet pas de distinguer les deux processus. Cependant, l'estimation globale de la distribution verticale du flux d'apport de carbone aux matières organiques peut être tirée de l'équation 1. Nous postulons que la répartition du carbone nouveau au temps t = 0 est une estimation de la distribution verticale du flux d'apport de carbone de la végétation aux matières organiques du sol.
28Dans l'ensemble maïs + Miscanthus, ces apports sont répartis en 81 % dans la couche 0-30 cm et 19 % dans la couche 30-100 cm, avec une erreur standard de ± 4 %.
29Par le même calcul de régression, nous pouvons représenter la répartition de ces apports par couche de 10 cm (Figure 4). Les couches inférieures plus profondes que 40 cm ont été regroupées par ensembles de 20 cm de profondeur dans la présentation pour limiter la dispersion statistique. À l'intérieur de la couche 0-30 cm, la distribution des apports n'est pas uniforme, contrairement à la répartition du carbone total du sol (Figure 2), car la population analysée comprend plusieurs sites non labourés depuis la mise en place de la monoculture : cultures pérennes et conduites en travail du sol réduit.
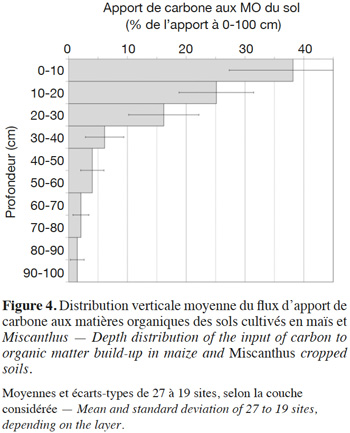
4. Discussion
30Il s'agit à notre connaissance d'une première synthèse des mesures de renouvellement du carbone profond sur un tel nombre de sites. Notre étude ne s'appuie que sur quelques cultures et ne peut être extrapolée directement à tous les sites cultivés. Outre les variations d'allocation de carbone souterrain et de développement des systèmes radiculaires, il est attendu que la proportion relative de carbone apporté en profondeur soit d'abord dépendante du régime de restitution des parties aériennes en surface. Dans notre jeu de données, les restitutions sont celles pratiquées le plus classiquement, c'est-à-dire restitution des tiges et feuilles de maïs, restitutions obligatoires à la coupe des Miscanthus. Concernant la canne à sucre, les pratiques ont pu être différentes, avec du brûlage des feuilles fréquemment pratiqué par le passé. Concernant l’ensemble maïs et Miscanthus, nous n'avons pas d'indication que l'allocation souterraine soit significativement différente de celle des céréales, ni même des poacées cultivées en fourrage. En revanche, les légumineuses sont connues pour leur enracinement profond et leur contribution significative au carbone profond a été établie (Guan et al., 2015).
31Dans un objectif de modélisation du bilan organique des sols, on pourrait ajouter au bilan du C superficiel celui du C profond, comme le proposent Jenkinson et al. (2008). Le modèle AMG (Andriulo et al., 1999 ; Bouthier et al., 2014) est l’un des modèles les plus simples du renouvellement du carbone utilisé en agriculture. Le carbone est réparti en un compartiment actif recevant une proportion k1 des apports annuels m et une constante de minéralisation k2. À ce carbone actif s'ajoute un compartiment CS de C stable à l'échelle du siècle. On peut ajouter un compartiment de C actif profond, mais même dans ce cadre très simplificateur, il peut y avoir plusieurs stratégies : représenter ou non un flux de transfert du carbone actif de la surface vers la profondeur, considérer les vitesses de biodégradations k2 comme égales ou non en surface et en profondeur, estimer a priori le C stable profond, etc. Nous proposons pragmatiquement la représentation la plus simple : vitesses de biodégradation identiques et transport non représenté explicitement. Sur la base des données à moyen terme (20 ans) du Miscanthus et du maïs, cette proposition revient :
32– à considérer un compartiment de carbone actif 30-100 cm égal à 0,30 (± 0,07, 1σ) fois le compartiment actif de la couche 0-30 cm, le reste étant du C stable,
33– à ajouter, dans les systèmes avec restitution des résidus de récolte dits « facultatifs », au flux d'apport à la couche 0-30, un apport à la couche 30-100 cm égal aussi à 0,30 fois celui alloué à la couche 0-30 cm,
34– d'appliquer en profondeur les mêmes paramètres (k1, k2) de la dynamique du compartiment dit « actif » du modèle qu'en surface.
35Le transport vertical de carbone est ici intégré implicitement sur 20 ans dans l'ensemble k1m, k2. La proportion allouée en profondeur est estimée dans les systèmes avec restitution des parties aériennes dites « facultatives » (tiges et feuilles ou feuilles sénescentes). Il s'agit bien sûr d'une approximation simplificatrice dont on sait qu'elle est probablement biaisée, en raison des vitesses de biodégradation plus lentes en profondeur, du transport vertical de carbone qui peut être très variable et des effets de priming (Fontaine et al., 2007), que l'équation linéaire de minéralisation d'ordre 1 ignore. Un effort de recherche important doit être mené pour paramétrer ces processus.
36La variance indiquée ci-dessus montre une très grande incertitude dans l'estimation et donc un risque d'erreur à 95 % sur une estimation isolée proche de la moitié du flux lui-même. La variabilité réelle (disparité entre sites) est certainement plus faible que la variabilité observée à partir de notre étude. En effet, une part de la variance vient de la méthode elle-même, notamment de la limite de sensibilité du traçage isotopique. D'autre part, une difficulté récurrente dans l'analyse des profils de sols publiés concerne l'information à la limite inférieure des profils : la plupart des auteurs n'indiquant pas l'épaisseur du sol étudié jusqu'à un horizon de roche mère, si bien qu'il y a un risque de confusion entre sol non échantillonné et absence de terre (concentration en carbone nulle) dans les publications. Nous recommandons de spécifier systématiquement l'épaisseur naturelle du sol et, si possible, d'apporter une description pédologique même sommaire des horizons profonds. Il est certain que les modèles globaux, comme régionaux, de la dynamique du carbone devront bénéficier d'informations détaillées, à la fois sur la profondeur des sols, les caractéristiques pédologiques et les stocks de carbone, comme le propose l'initiative mondiale GlobalSoilMap (Arrouays et al., 2015).
37Les valeurs de proportion du carbone incorporé dans les différentes couches que nous rapportons doivent être interprétées en prenant en compte les différents mécanismes contribuant à l'apport de carbone en profondeur. En tout premier lieu et de façon triviale, en cas de labour plus profond que 30 cm, le bilan de carbone doit a minima prendre en compte l'ensemble de la couche homogénéisée pour les bilans de C. Les autres apports sont le transport naturel par bioturbation ou lixiviation et les apports racinaires, les deux pouvant agir en interaction, via le comblement des pores racinaires ou leur colonisation par la faune du sol. Les proportions de carbone alloué aux matières organiques profondes observées ici sont plus importantes que les estimations de biomasse souterraines habituellement rapportées, confirmant l'observation générale qu’à biomasse équivalente, les racines contribuent plus aux matières organiques des sols que les tiges et les feuilles (Balesdent et al., 1996 ; Rasse et al., 2005). Les causes évoquées sont nombreuses, en particulier la contribution des exsudats et rhizodépôts, et la préservation plus importante du carbone issu des tissus racinaires, des composés solubles ou des métabolites microbiens qui en sont issus, en interaction possible avec la sorption et la protection par les phases minérales.
5. Conclusions
38Les matières organiques situées en dessous de la couche arable contiennent une partie non négligeable du carbone actif des sols. Nous concluons que ces matières organiques profondes ne doivent être négligées ni dans les bilans de carbone, ni dans l'estimation de la fourniture d'azote par minéralisation des matières organiques. Les rapports C/N, systématiquement plus petits en profondeur qu'en surface dans les sols, indiquent que les matières organiques profondes sont très azotées et que leur renouvellement dans les couches profondes peut libérer de l'azote minéral dans des proportions proches de celles des flux de carbone, et devrait donc également être pris en compte dans les bilans d'azote.
39Dans un objectif de comptabilité du carbone en agriculture, pour le bilan carbone ou pour la gestion de la culture, la prise en compte des variations du C profond est donc nécessaire. Cependant, l'estimation de variations temporelles par mesure directe au champ fait face à des difficultés énormes, largement supérieures à celles rencontrées pour le carbone superficiel, en raison notamment de la variabilité spatiale intra-parcellaire généralement forte, de la lenteur des changements et de l'imprécision sur la mesure des concentrations faibles en carbone (quelques mg·g-1). Les estimations des évolutions temporelles du stock de carbone profond des sols et de leurs facteurs de variation devront donc s'appuyer sur les essais agronomiques de longue durée particulièrement bien contrôlés ou sur la modélisation calée sur des sites fortement instrumentés. Elles devraient être systématiquement accompagnées de descriptions pédologiques pour faciliter leur interprétation. Favoriser les apports de carbone par les systèmes racinaires pour augmenter le stockage de carbone en profondeur est une technique qui mérite attention. Il pourrait s'agir de choix d'espèces ou de génotypes intraspécifiques, prenant en compte également l'interaction entre les génotypes et l'environnement tellurique et pédoclimatique. Il faudra cependant estimer de façon concomitante les conséquences potentielles de l'effet de sur-minéralisation (priming effect) des matières organiques anciennes du fait de l’apport aux micro-organismes du sol d’exsudats racinaires riches en énergie.
40Remerciements
41Cette étude a été partiellement financée par le projet ANR « Dedycas » (Gestion Sobre des Ressources 14-CE01-0004). Nous remercions Christine Marol, Sylvie Milin, Patrick Signoret et Samuel Legros pour leur contribution aux analyses isotopiques additionnelles et plusieurs scientifiques qui nous ont élégamment fourni les données numériques de leurs données publiées.
Bibliographie
Ahlström A., Schurgers G., Arneth A. & Smith B., 2012. Robustness and uncertainty in terrestrial ecosystem carbon response to CMIP5 climate change projections. Environ. Res. Lett., 7, 044008.
Alexander E.B., 1980. Bulk densities of California soils in relation to other soil properties. Soil Sci. Soc. Am. J., 44, 689-692.
Andriulo A., Mary B. & Guérif J., 1999. Modelling soil carbon dynamics with various cropping sequences on the rolling pampas. Agron. EDP Sci., 19(5), 365-377.
Arrouays D. et al., 2015. GlobalSoilMap: towards a fine-resolution global grid of soil properties. Adv. Agron., 125, 93-134.
Balesdent J. & Balabane M., 1996. Major contribution of roots to soil carbon storage inferred from maize cultivated soils. Soil Biol. Biochem., 28, 1261-1263.
Balesdent J. et al., 2011. Contribution de la rhizodéposition aux matières organiques du sol, quelques implications pour la modélisation de la dynamique du carbone. Étude Gestion Sols, 18(3), 201-216.
Balesdent J. & Mariotti A., 1996. Measurement of soil organic matter turnover using 13C natural abundances. In: Boutton T.W & Yamasaki S.I., eds. Mass spectrometry of soils. New York, USA: Marcel Dekker Inc., 83-111.
Bouthier A. et al., 2014. Adaptation et mise en oeuvre du modèle de calcul de bilan humique à long terme AMG dans une large gamme de systèmes de grandes cultures et de polyculture-élevage. Innovations Agron., 34, 125-139.
Carvalhais N. et al., 2014. Global covariation of carbon turnover times with climate in terrestrial ecosystems. Nature, 514, 213-217.
Ciais P. et al., 2013. Carbon and other biogeochemical cycles. In: Stocker T.F. et al., eds. Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 465-570.
Cerri C. et al., 1985. Application du traçage isotopique naturel en 13C à l'étude de la dynamique de la matière organique dans les sols. C. R. Acad. Sc., Ser. II, 9, 423-428.
Coleman K. & Jenkinson D.S., 1995. RothC-26 3. A model for the turnover of carbon in soil: model description and users guide. Harpenden, UK: Rothamsted Research.
Elzein A. & Balesdent J., 1995. Mechanistic simulation of vertical-distribution of carbon concentrations and residence times in soils. Soil Sci. Soc. Am. J., 59, 1328-1335.
Fontaine S. et al., 2007. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature, 450, 277-280.
Guan X.-K. et al., 2016. Soil carbon sequestration by three perennial legume pastures is greater in deeper soil layers than in the surface soil. Biogeosciences, 3, 527-536.
Guenet B. et al., 2013. The relative importance of decomposition and transport mechanisms in accounting for soil organic carbon profiles. Biogeosciences, 10, 2379-2392.
Hansson P.J., Edwards N.T., Garten C.T. & Andrews J.A., 2000. Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry, 48, 115-146.
Harper R.J. & Tibbett M., 2013. The hidden organic carbon in deep mineral soils. Plant Soil, 368, 641-648.
Hiederer R. & Köchy M., 2011. Global soil organic carbon estimates and the harmonized world soil database. EUR 25225 EN. Luxembourg: Publications Office of the European Union.
IUSS Working Group WRB, 2014. World Reference Base for Soil Resources 2014. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. Roma: FAO.
Jenkinson D.S. & Coleman K., 2008. The turnover of organic carbon in subsoils. Part 2. Modelling carbon turnover. Eur. J. Soil Sci., 59, 400-413.
Jobbagy E.G. & Jackson R.B., 2000. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl., 10, 423-436.
Koven C.D. et al., 2011. Permafrost carbon-climate feedbacks accelerate global warming. Proc. Natl. Acad. Sci. U.S.A., 108, 14769-14774.
Mathieu J., Hatté C., Parent E. & Balesdent J., 2015. Deep soil carbon dynamics are driven more by soil type than by climate: a worldwide meta-analysis of radiocarbon profiles. Global Change Biol., 21, 4278-4292.
Mulder V.L. et al., 2016. National versus global modelling the 3D distribution of soil organic carbon in mainland France. Geoderma, 26, 16-34.
Rasse D.P., Rumpel C. & Dignac M.F., 2005. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil, 269, 341-356.
Rumpel C. & Kögel-Knabner I., 2011. Deep soil organic matter—a key but poorly understood component of terrestrial C cycle. Plant Soil, 338,143-158.
Salomé C. et al., 2010. Carbon dynamics in topsoil and in subsoil may be controlled by different regulatory mechanisms. Global Change Biol., 16, 416-426.

