


- Accueil
- Volume 20 (2016)
- Numéro 1
- Mesure des flux de CO2 et séquestration de carbone dans les écosystèmes terrestres ouest-africains (synthèse bibliographique)


Visualisation(s): 2526 (37 ULiège)
Téléchargement(s): 410 (4 ULiège)

Mesure des flux de CO2 et séquestration de carbone dans les écosystèmes terrestres ouest-africains (synthèse bibliographique)

Notes de la rédaction
Reçu le 2 mars 2015, accepté le 9 octobre 2015
Résumé
Introduction. En Afrique de l’Ouest comme ailleurs dans d’autres régions du monde, la préservation des écosystèmes terrestres s’avère primordiale pour toute politique d’atténuation des émissions de gaz à effet de serre, en particulier du dioxyde de carbone. Une évaluation pertinente de la contribution d’une région donnée au cycle global du carbone implique la nécessité de raisonner en termes de bilan en considérant dans l’analyse tous les flux d’exportation et d’importation dans les écosystèmes.
Littérature. Une synthèse bibliographique, basée sur les résultats publiés ou non de quelques études ayant porté sur les écosystèmes ouest-africains, a été conduite avec pour objectif principal d’évaluer l’importance des échanges de carbone avec l’atmosphère. Il ressort que les flux de carbone présentent une importante variabilité spatiale, principalement liée à la variabilité des conditions pédo-climatiques, des modes de gestion, des types de végétation et des formes d’usage des écosystèmes. La pluviométrie moyenne annuelle apparait être le facteur principal contrôlant les échanges de carbone dans la région. La variabilité spatiale de la photosynthèse explique 71 % de celle de cumuls annuels de NEE. Cet article confirme le comportement de puits de carbone de l’ensemble des écosystèmes terrestres ouest-africains considérés avec une séquestration moyenne annuelle de -2,0 ± 1,5 t C·ha-1 et fait apparaitre la nécessité d’améliorer la compréhension des mécanismes ou des facteurs qui contrôlent les échanges de carbone entre les écosystèmes et l’atmosphère.
Conclusions. Considérant les résultats de quelques sites, il apparait nécessaire de poursuivre les mesures des flux de carbone à l’échelle de l’écosystème sur le long terme afin de mieux apprécier leurs variabilités spatio-temporelles, notamment saisonnières et inter-annuelles. Cet article met également en évidence la nécessité d’étendre les mesures d’eddy covariance dans la région et à d’autres types de végétation de manière à faciliter des études de comparaison entre différents types d’écosystème. Ces études permettront de suggérer des modes de gestion plus durables favorisant la séquestration de carbone tout en limitant les émissions.
Abstract
CO2 measurement and carbon sequestration in West African terrestrial ecosystems. A review
Introduction. In West Africa, as elsewhere in other parts of the world, preserving terrestrial ecosystems has proven essential to any emissions mitigation policy for greenhouse gases, notably carbon dioxide. A meaningful assessment of the contribution of a given region to the global carbon cycle implies the need to think in terms of balance, taking into account the totality of the carbon export and import within the ecosystems.
Literature. This review, based on the results of a small number of published and unpublished studies focusing on West African ecosystems, has as its main objective the assessment of the importance of carbon exchange. Carbon fluxes appear to be spatially highly variable in the West African region, mainly due to the variability of the types of vegetation, approaches to ecosystem management, and edaphic and climatic conditions. The mean annual precipitation appears to be the main factor controlling plant respiration and photosynthesis in this region. Annual cumulated NEE has been positively correlated with the rate of photosynthesis, and has been shown to account for 71% of its spatial variability at the annual scale. This paper also confirms the presence of carbon sink behavior for considered ecosystems in the West Africa with an annual average carbon sequestration of -2.0 ± 1.5 t C·ha-1 and highlights the need to improve understanding of factors or mechanisms controlling the carbon exchange between the ecosystems and the atmosphere.
Conclusions. Based on these results, which were collected on a very limited number of sites, it appears necessary to continue with the measurement of carbon fluxes in the long term at the ecosystem scale in order to better appreciate the temporal and spatial variabilities, especially on a seasonal and inter-annual basis. This paper highlights the need to extend eddy covariance measurements to other vegetation types in this region in order to facilitate studies comparing the different ecosystems. This could be used as a basis for the recommendation of more sustainable management strategies favoring carbon sequestration while limiting emission levels.
Table des matières
1. Introduction
1Bien que les impacts négatifs associés aux émissions de CO2 soient clairement reconnus et identifiés (Houghton et al., 2012 ; GIEC, 2013 ; Grace et al., 2014 ; Mbow, 2014), il est fort probable que dans les prochaines décennies ces émissions se poursuivront, voire s’intensifieront. Dans le bilan global de carbone, les écosystèmes terrestres sont reconnus comme jouant un rôle essentiel, absorbant près de 30 % du CO2 anthropique total émis (GIEC, 2013). Toutefois, ces puits cachent une grande variabilité spatiale et inter-annuelle qui est mal comprise. Dans ce contexte, les écosystèmes terrestres africains demeurent jusqu’à présent très peu étudiés, comparés à ceux d’autres régions du monde (Mbow, 2014 ; Valentini et al., 2014).
2En termes d’émission de carbone, la contribution du continent africain aux émissions anthropiques totales de la planète est la plus faible, de l’ordre de 7 % du global. En revanche, le continent africain contribue pour 17 % à l’ensemble des émissions de la planète issues des changements dans les formes d’utilisation et les modes de gestion des sols (Canadell et al., 2009). Les modifications des modes d’usage des terres contribuent pour 48 % aux émissions totales de carbone de l’Afrique. Les émissions des écosystèmes africains sont aussi caractérisées par une grande variabilité spatiale et inter-annuelle (Ciais et al., 2011 ; Houghton et al., 2012 ; Valentini et al., 2014). L’Afrique de l’Ouest, qui a connu une grande et longue crise de sècheresse sans précédent dans les années 1970 et 1980 (Ago et al., 2005 ; Ozer et al., 2010 ; Fisher et al., 2013 ; Nicholson, 2013), est caractérisée par une forte variabilité climatique et un taux de croissance démographique très élevé (> 3 %) (Nicholson, 2013 ; UNDP, 2015). Cette région est par ailleurs soumise à divers processus lents de dégradation environnementale comme la désertification (Wittig et al., 2007 ; Ozer et al., 2010). Tous ces facteurs ont eu sans doute des impacts significatifs sur la croissance végétative et les flux de carbone dans les écosystèmes (Bombelli et al., 2009 ; Gonzalez et al., 2012 ; Abdi et al., 2014 ; Ajonina et al., 2014 ; Valentini et al., 2014). Dans ce contexte, il s’avère impérieux de mieux comprendre la dynamique de la végétation et des échanges de carbone, et de les quantifier. De façon spécifique, il s’agira d’étudier les réponses des flux de carbone aux facteurs pédo-climatiques et aux modes de gestion des écosystèmes. Ainsi, il sera possible de mieux appréhender la variabilité spatiale et temporelle des échanges de carbone, puis d’estimer les bilans carbonés des principaux écosystèmes africains. Ceci contribuera à réduire significativement les larges incertitudes jusqu’à présent observées sur les flux de carbone dans cette région. Il sera possible de mieux prédire l’évolution des échanges de carbone entre ces écosystèmes et l’atmosphère en tenant compte des changements climatiques et des évènements météorologiques extrêmes. Ainsi, il sera possible d’émettre à long terme des suggestions pertinentes sur les modes de gestion plus durables des écosystèmes africains. La contribution au cycle global de carbone de l’Afrique de l’Ouest pourra être mieux estimée. In fine, les vulnérabilités des écosystèmes pourront également être mieux déterminées et évaluées.
3L’objectif principal de cet article est de faire l’état des connaissances sur les échanges de carbone dans les écosystèmes terrestres ouest-africains. Une synthèse bibliographique et de résultats expérimentaux non publiés sera réalisée afin de faire le bilan des connaissances sur les flux de carbone en Afrique de l’Ouest. Des recherches et des mesures complémentaires nécessaires à une connaissance approfondie des échanges de carbone, de leurs variabilités aux différentes échelles spatio-temporelles et du fonctionnement des écosystèmes terrestres seront finalement discutées.
2. L’Afrique de l’Ouest, ses spécificités climatiques et les sites d’étude considérés
4L’Afrique de l’Ouest est la région située au Sud du Sahara comprise entre 18°W et 15°E s’étendant de 4°N jusqu’à 25°N (Figure 1). Elle jouit de trois principaux types de climats prédominants définis par Le Lay (2006) sur la base de la moyenne annuelle de précipitation (MAP) : le climat tropical humide ou guinéen (MAP > 1 400 mm), le climat soudanien ou tropical sec (700 < MAP < 1 400 mm) et le climat sahélien (MAP < 700 mm). À ce gradient pluviométrique Nord-Sud, caractérisé par une saisonnalité très marquée dans chacune des zones, est associé un gradient de grands ensembles de végétation, laquelle est globalement constituée de forêts, de savanes, de mangroves, de pâturages, de jachères, de cultures et de plantations (Aubréville, 1949 ; Kaptue et al., 2010 ; Abdi et al., 2014). En 2010, la population de cette région était estimée à 305 millions d’habitants avec une densité moyenne de 50 habitants par km2 (UNDP, 2015).
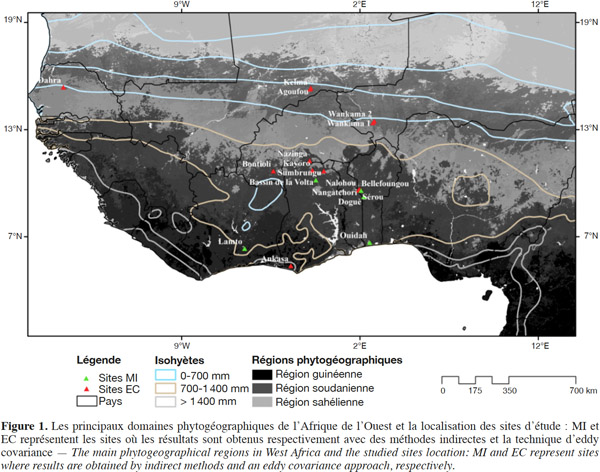
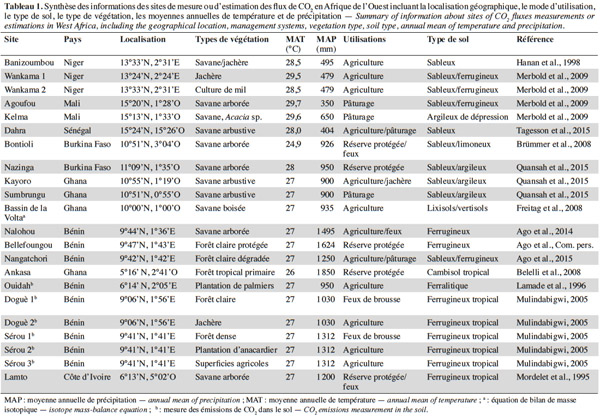
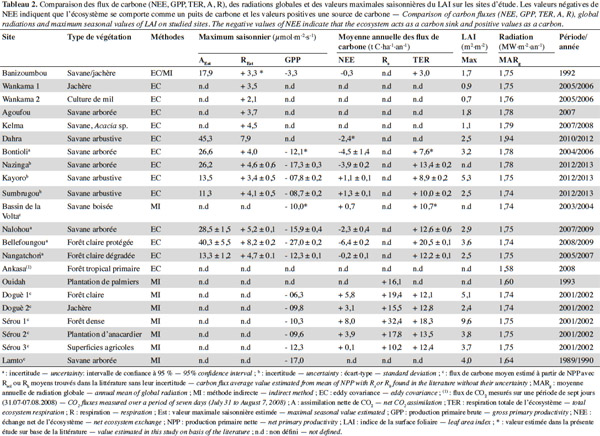
5Les résultats présentés dans cet article concernent des écosystèmes répartis sur trois grandes régions phytogéographiques (Figure 1), 18 localités et 22 sites (Tableau 1). Neuf sites de mesure des flux de carbone ont été installés dans le cadre de programmes et projets internationaux comme AMMA-CATCH et CarboAfrica qui ont par ailleurs établi une étroite collaboration avec le réseau mondial FluxNet (Merbold et al., 2009 ; Sjöström et al., 2013 ; Ago et al., 2014 ; Valentini et al., 2014 ; Ago et al., 2015 ; FluxNet, 2015 ; Quansah et al., 2015). Treize autres sites ont été mis en place dans diverses études et campagnes de mesure isolées supportées par des projets d’envergure nationale. En général, la période de mesure des flux de carbone varie de quelques mois à plus de deux années pour la majorité des sites, excepté la forêt tropicale primaire d’Ankasa au Ghana pour laquelle les flux nets de CO2 concernent seulement une semaine (Tableau 2). La première campagne de mesures des flux nets de CO2 a été effectuée en 1992 à Banizoumbou (savane/jachère) au Niger. Les plus longues séries de mesures disponibles actuellement concernent la période de 2004 à 2012 et les sites de Dahra (savane) au Sénégal, de Bontioli (savane) au Burkina Faso, de Nalohou (savane), de Bellefoungou et de Nangatchori (forêt claire) au Bénin. Les mesures des flux de carbone sont toujours réalisées jusqu’à présent sur les sites de Bellefoungou et de Nalohou au Bénin.
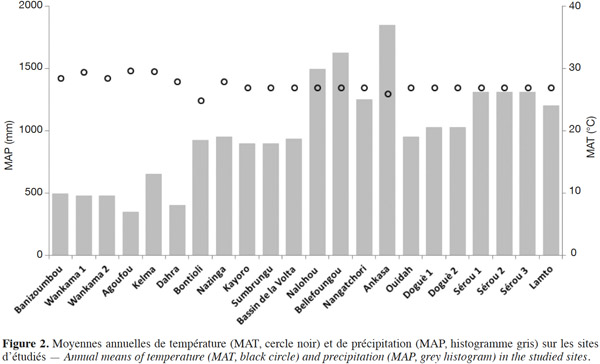
6Sur les 22 sites considérés, dont 14 sont équipés d’un système d’eddy covariance, la durée totale de la saison sèche varie globalement de cinq mois dans la région guinéenne (Ankasa au Ghana) à huit mois environ au Sahel (Agoufou au Mali). Les sites sous climat tropical humide sont caractérisés par la succession annuelle de quatre saisons : deux sèches et deux pluvieuses en alternance et d’inégales durées induites par les déplacements de la Zone de Convergence Inter Tropicale (ZCIT). Les sites soudano-sahéliens sont caractérisés par une seule saison sèche et une seule saison pluvieuse d’inégales durées. Les moyennes annuelles de précipitation (MAP) évoluent également en fonction de la localisation des sites (Figure 1, Tableau 1), le Sahel étant la région la moins arrosée et la région guinéenne la plus arrosée (Le Lay, 2006). Pour l’ensemble des sites, les MAP se situent entre 350 mm (Agoufou au Mali) et 1 850 mm (Ankasa au Ghana). Les moyennes annuelles de température (MAT) varient de 24 à 30 °C, avec les plus faibles valeurs enregistrées dans la région guinéenne et les plus élevées dans le Sahel (Figure 2, Tableau 1).
3. Cycle de carbone à l’échelle de l’écosystème et méthodes de mesure des différents composants
7Afin de mieux décrire les méthodes de mesure et d’estimation des échanges nets de carbone, il est très important de rappeler quelques définitions relatives aux divers échanges de CO2 dans les écosystèmes. La production primaire brute (GPP : gross primary productivity) représente le CO2 total prélevé par les végétaux chlorophylliens de l’atmosphère via la photosynthèse. La respiration totale de l’écosystème (TER : total ecosystem respiration) désigne la somme des respirations autotrophe (Ra) (CO2 rejeté par la plante dans l’atmosphère) et hétérotrophe (Rh) (émission du CO2 provenant de la décomposition de la matière organique). Quant à l’échange net (NEE : net ecosystem exchange), il désigne la somme de la GPP et de la TER. Enfin, la production primaire nette (NPP : net primary productivity) quantifie le carbone stocké dans les différentes parties de la végétation.
8La technique d’eddy covariance, dont les bases théoriques sont décrites en détail par Aubinet et al. (2012), permet la mesure directe de l’échange net de CO2 (NEE) entre l’écosystème et l’atmosphère. Cette méthode est utilisée depuis 1993 par plusieurs programmes et projets régionaux principalement EuroFlux-CarboEurope, AmeriFlux, AsiaFlux, CarboAfrica et AMMA-CATCH qui ont été fédérés à l’échelle mondiale dans le réseau FluxNet. Celui-ci regroupait, en février 2015, au total 679 sites dont 25 seulement étaient installés dans des écosystèmes africains, soit 4 % environ (FluxNet, 2015). En Afrique de l’Ouest, seulement 14 stations de mesure de flux de carbone étaient installées sur différents types de végétation (Figure1, Tableau 1). Elles étaient réparties dans cinq pays à savoir le Bénin (3), le Burkina Faso (2), le Ghana (3), le Mali (2), le Niger (3) et le Sénégal (1). La plupart des stations ont permis de réaliser des campagnes de mesures de flux de carbone de façon temporaire ou saisonnière, mais rarement de façon continue à long terme à cause des couts très élevés des capteurs nécessaires, des difficultés d’installation d’un système d’eddy covariance, de leur maintenance, et des exigences liées aux analyses et aux traitements des données notamment l’évaluation des données manquantes (Aubinet et al., 2012) pour l’établissement du bilan carboné des écosystèmes. Malgré ces difficultés de mesure en continu des flux et les exigences de traitement, cette technique demeure actuellement la plus répandue dans le monde (FluxNet, 2015). Elle fournit une évaluation des échanges nets de gaz entre les écosystèmes terrestres et l’atmosphère à une échelle spatiale de quelques hectares autour du point de mesure. Elle nécessite néanmoins que l’écosystème soit raisonnablement homogène et elle fournit des mesures qui sont spécifiques au site exploré. Une extrapolation spatiale nécessite donc un travail en réseau et une multiplication des sites de mesures afin de pouvoir comparer les résultats (Aubinet et al., 2012). Ces différentes mesures peuvent être sommées aux échelles horaire, journalière, mensuelle, annuelle et pluri-annuelle. En raison de sa haute résolution temporelle, elle est adaptée pour étudier la physiologie des écosystèmes. Enfin, cette méthode, en fournissant des mesures de NEE, permet une évaluation de la séquestration ou du rejet du carbone par un écosystème mais ne permet pas, sans traitement de données supplémentaires, d’analyser de manière fine les mécanismes responsables des variabilités de la GPP et de la TER.
9Par ailleurs, d’autres méthodes indirectes permettent également d’approcher la NEE en utilisant la relation ci-dessous :
10NEE = NPP + Rh (1)
11Ces méthodes nécessitent une estimation des deux variables, à savoir NPP (comptée négativement) et Rh (comptée positivement). La NPP est estimée à partir de l’accroissement de biomasse en utilisant des relations allométriques établies sur la base des quantités de litière ou des résultats d’inventaires en considérant certaines espèces spécifiques de l’écosystème (Bombelli et al., 2009 ; Ciais et al., 2011 ; Valentini et al., 2014). La NPP est exprimée en termes de quantité de carbone, admettant que la fraction en carbone de la biomasse est comprise entre 40 et 55 % (Schlesinger, 1997 ; Carvalho et al., 1998 ; Hanan et al., 1998 ; Malhi et al., 1999). La respiration hétérotrophe (Rh) est difficile à déterminer expérimentalement, notamment la séparation des deux composantes de la respiration du sol, Rh et Ra. L’interprétation des mesures est aussi délicate à cause des interactions complexes entre les différents pools de carbone dans le sol et les micro-organismes (Hanson et al., 2000 ; Bond-Lamberty et al., 2004). Afin de contourner cette difficulté majeure, Rh est estimée à partir des mesures de la respiration du sol (Rs) grâce à l’équation établie par Bond-Lamberty et al. (2004) :
12Ln (Rh) = 1,22 + 0,73 Ln (Rs) (2)
13avec Rh et Rs exprimés en g C·m-2·an-1. L’équation est très hautement significative avec 0,81 comme coefficient de détermination. Cette équation fut choisie car elle a été établie sur la base des mesures effectuées sur 53 sites répartis dans le monde (y compris la région tropicale). Aucun effet significatif de la méthode de mesure, du type de biome, de la localisation géographique du site, du type de climat et des propriétés hydrologiques du sol n’a été trouvé.
14Dans le présent article, cette relation (2) nous a permis d’estimer la Rh des sites de Sérou et de Doguè localisés au Nord-Ouest du Bénin en utilisant les mesures de respiration du sol (Rs) réalisées pendant la période 2001-2002 par Mulindabigwi (2005) avec un analyseur de gaz infrarouge de type EGM-2 connecté à une chambre de respiration du sol SRC-1 (PP Systems, Haverhill, MA).
15Pour les sites ne disposant pas d’un système d’eddy covariance mais dont la NPP a déjà été estimée par d’autres auteurs, nous avons déterminé la GPP par la relation (3) ci-dessous où α désigne l’efficience d’utilisation du carbone par l’écosystème considéré. Dans la région tropicale, α varie entre 0,40 à 0,53 à l’échelle annuelle (Landsberg et al., 1997 ; Hanan et al., 1998 ; Malhi et al., 1999 ; DeLucia et al., 2007 ; Luyssaert et al., 2007) avec une valeur moyenne égale à 0,43 et un intervalle de confiance de 0,10. Cette valeur moyenne de α a été utilisée pour déterminer la GPP des sites de Lamto en Côte d’Ivoire, du bassin de la Volta au Ghana, de Sérou et de Doguè au Bénin (Tableau 2) grâce aux valeurs de NPP de ces sites de la littérature (Mordelet et al., 1995 ; Mulindabigwi, 2005 ; Freitag et al., 2008).
16GPP = 1/α*NPP (3)
17Finalement, la TER des écosystèmes ne disposant pas de système d’eddy covariance a été déterminée en additionnant les respirations autotrophe et hétérotrophe. La respiration autotrophe a été estimée par la relation ci-dessous (Schulze, 2006 ; Luyssaert et al., 2007) :
18Ra = NPP - GPP (4)
19Globalement, l’estimation de la NEE à partir des méthodes indirectes pour les écosystèmes africains apparait souvent difficile à cause des contraintes d’échantillonnage représentatif de l’écosystème, des larges incertitudes observées sur les résultats finaux, de l’importance de la main-d’œuvre nécessaire et du caractère statique des données d’inventaire (Bombelli et al., 2009 ; Ciais et al., 2011 ; Fisher et al., 2013 ; Mbow, 2014 ; Valentini et al., 2014). Ces approches indirectes ont permis notamment d’avoir des estimations des stocks de carbone des écosystèmes, mais pas d’analyser les flux de carbone, leurs dynamiques et les facteurs ou mécanismes qui contrôlent ceux-ci.
20L’un des facteurs majeurs permettant d’apprécier la productivité d’une végétation ou d’un écosystème est son indice foliaire (Leaf Area Index pour les anglophones, abrégé par LAI), c’est-à-dire la surface des feuilles exprimée par unité de surface du sol. Le LAI peut être estimé à travers plusieurs méthodes. Dans la présente étude, les valeurs de LAI utilisées proviennent des campagnes de mesures optiques directes, des mesures de la litière foliaire au sol et des estimations basées sur des données du satellite SEVIRI contraintes par les mesures in situ faites par photographie hémisphérique. Le LAImax exprime le maximum saisonnier annuel, c’est-à-dire la valeur maximale de LAI durant la saison humide. Les moyennes annuelles de radiation globale (MARg) proviennent également des estimations basées sur des données satellitaires (http://www.soda-is.com).
21En somme, une combinaison d’approches d’inventaire et d’approches micro-météorologiques pour les mesures des flux de carbone dans les écosystèmes terrestres ouest-africains peut être mobilisée dans la perspective de réduire les incertitudes sur les résultats de mesures ou d’estimations des flux.
4. Évolution des flux de carbone dans les écosystèmes terrestres ouest-africains
22La pluviométrie constitue à l’échelle saisonnière l’un des principaux facteurs climatiques contrôlant la dynamique des flux de carbone dans les écosystèmes terrestres (Brümmer et al., 2008 ; Williams et al., 2008 ; Ciais et al., 2009 ; Merbold et al., 2009 ; Jung et al., 2011 ; Fisher et al., 2013 ; Ago et al., 2014 ; Ago et al., 2015 ; Quansah et al., 2015 ; Tagesson et al., 2015) et la dynamique du couvert végétal (Hiernaux et al., 2009 ; Seghieri et al., 2009) de cette région caractérisée par l’alternance entre saisons pluvieuses et saisons sèches, toutes deux d’inégales durées. De plus, plus que le volume total d’eau, c’est surtout la régularité et la répartition des pluies, la durée totale de la période humide et l’humidité du sol qui importent (Seghieri et al., 2009).
23Les principales variables climatiques influençant les dynamiques des écosystèmes terrestres et celles des flux de carbone (GPP, TER et NEE) sont le rayonnement, la pluviométrie et la température (Nemani et al., 2003 ; Garbulsky et al., 2010 ; Aubinet et al., 2012 ; Sjöström et al., 2013). Les cumuls annuels des flux de carbone TER et GPP en fonction des variables climatiques sont représentés respectivement aux figures 3 et 4. On observe la plus faible valeur de la TER (3,0 t C·ha-1·an-1) à Banizoumbou (savane/jachère) au Niger et la plus élevée (20,5 t·ha-1·an-1) à Bellefoungou (forêt claire protégée) au Bénin, avec comme valeur moyenne 12,0 ± 2,3 t·ha-1·an-1 en considérant l’ensemble des sites.
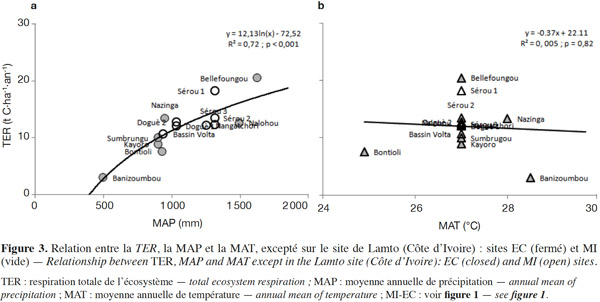
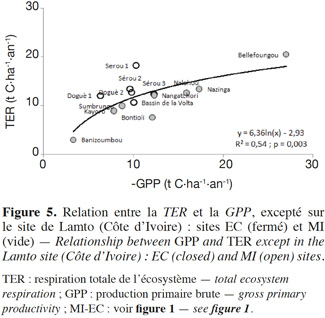
24Aucune corrélation significative n’est trouvée entre la TER et la MAT (Figure 3b), par contre une tendance pluviométrique claire apparait dans la variation spatiale de la TER (Figure 3a), ce qui est confirmé par la corrélation positive très hautement significative (R2 = 0,72 et p < 0,001) entre la TER et la MAP. Ceci suggère le contrôle de la TER par les précipitations annuelles en Afrique de l’Ouest. La TER est limitée sur les sites recevant des précipitations annuelles faibles. Cependant, la TER semble être indépendante des MAP pour les sites recevant plus de 1 000 mm de précipitation par an (sites localisés entre les latitudes 6° et 11°N), ce qui se traduit par la corrélation non significative (R2 = 0,37 et p = 0,111) entre la TER et la MAP lorsqu’on considère uniquement ces sites. Ceux-ci ont une MAT comprise entre 26 et 28 °C et montrent une grande variabilité spatiale de la TER avec des valeurs de la TER variant de 12,1 (Doguè 1) à 20,5 t C·ha-1·an-1 (Bellefoungou) et même différentes pour plusieurs sites recevant une pluviométrie moyenne identique (Sérou 1 et 2, par exemple). Ceci suggère l’intervention dans cette zone des facteurs autres que les moyennes de températures et de précipitations. Ces autres facteurs peuvent être le pédo-climat, le type de végétation, le degré de perturbation, le mode de gestion et la forme d’utilisation des écosystèmes. La variabilité spatiale de la TER dans les écosystèmes ouest-africains peut être également liée à la qualité et à la quantité de la matière organique, un facteur principal de la respiration hétérotrophe (Hanson et al., 2000 ; Chambers et al., 2004). Il existe en effet des différences entre les sites concernant les types de sol, les types de végétation, les formes d’utilisation et les modes de gestion (Tableau 1). Donc, les types de végétation, l’historique des sites, les types de sols, les formes d’utilisation et les modes de gestion des différents écosystèmes constituent probablement des facteurs importants contrôlant la variation spatiale de la TER dans cette région (Chambers et al., 2004). Une corrélation positive significative a également été trouvée entre la TER et la MAP dans la région soudano-sahélienne par d’autres auteurs en considérant l’ensemble des types d’écosystèmes (Ciais et al., 2009 ; Jung et al., 2011 ; Sjöström et al., 2013) et en Afrique du Sud pour la savane semi-aride (Archibald et al., 2009). À l’échelle annuelle, la variabilité de la MAT n’est donc pas un facteur important influençant la variation spatiale de la TER (Valentini et al., 2000 ; Reichstein et al., 2003 ; Jung et al., 2011). Il est possible que le manque de relation claire entre la TER et la MAT soit induit par la saisonnalité très marquée des périodes sèches sur les sites (Liu et al., 2006 ; Richardson et al., 2006 ; Curiel Yuste et al., 2007 ; Williams et al., 2009) ou encore par la faible amplitude des variations de la MAT entre les sites (< 4 °C). Dans la forêt tropicale humide en Amérique centrale, Loescher et al. (2003) ont partiellement expliqué le manque de relation claire entre la respiration et la température par la faible amplitude des variations de la température nocturne (< 9 °C). Par ailleurs, la dépendance claire de la TER à la MAP trouvée pour les écosystèmes terrestres ouest-africains peut résulter de deux effets possibles : l’humidification du sol active la respiration hétérotrophe, le « Birch effect » (Lamade et al., 1996 ; Miller et al., 2005 ; Mulindabigwi, 2005 ; Davidson et al., 2006 ; Liu et al., 2006 ; Jarvis et al., 2007) et les précipitations peuvent également affecter la TER en contribuant à l’accroissement de la respiration autotrophe par une plus grande croissance végétative (Friborg et al., 1997 ; Hanan et al., 1998 ; Archibald et al., 2009 ; Ciais et al., 2009 ; Jung et al., 2011), ce qui est corroboré par la corrélation hautement significative (R2 = 0,54 et p = 0,003) trouvée entre la TER et la GPP (Figure 5), suggérant également la dépendance de la respiration autotrophe à la productivité des écosystèmes.
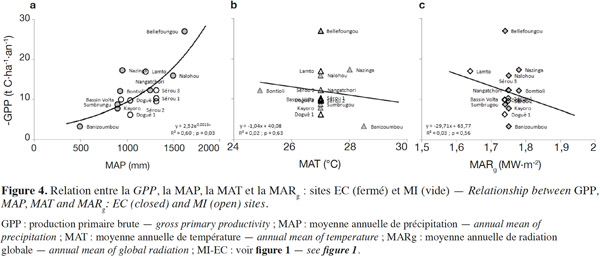
25Globalement, la GPP varie de -27,0 à Bellefoungou (forêt claire protégée) au Bénin à -3,3 t C·ha-1·an-1 sur le site de Banizoumbou (savane/jachère) au Niger (Tableau 2), avec une moyenne de -12,0 ± 2,9 t C·ha-1·an-1 si l’on considère tous les sites. Cette plage de variation de la GPP apparait très large a priori, mais recouvre parfaitement les valeurs reportées et comprises entre -40,0 et -0,5 t C·ha-1·an-1 pour une variété d’écosystèmes dans le monde (Valentini et al., 2000; Law et al., 2002 ; Luyssaert et al., 2007 ; Malhi, 2012 ; Sjöström et al., 2013 ; Grace et al., 2014 ; Valentini et al., 2014).
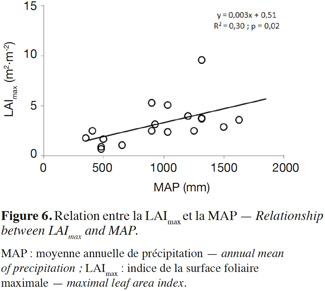
26Comme la TER, une corrélation significative (R2 = 0,60 et p = 0,03) est trouvée entre la GPP et la MAP, montrant que la variabilité spatiale de la GPP est clairement influencée par la MAP (Figure 4a). Aucune relation claire de la GPP avec la MAT et la MARg n’a été observée (Figure 4b et Figure 4c), suggérant que la variation de ces facteurs climatiques (température et radiation) entre les différents sites n’influence donc pas la variabilité spatiale de la GPP. Il est probable que la corrélation négative et très hautement significative (R2 = 0,50 et p < 0,001) trouvée entre la MAT et la MAP des sites pris en compte conduise à l’absence de l’expression de la MAT comme un facteur influençant la variabilité spatiale de la GPP. L’amplitude annuelle des variations de la MARg entre les différents sites est faible et pourrait expliquer l’absence d’une influence claire de ce facteur climatique sur la variabilité spatiale de la GPP. Plusieurs études à une plus large échelle spatiale ont reporté une diversité de valeurs de la GPP selon différents facteurs abiotiques et biotiques (Valentini et al., 2000 ; Law et al., 2002 ; Luyssaert et al., 2007 ; Merbold et al., 2009 ; Garbulsky et al., 2010). Les valeurs de GPP les plus élevées (en valeur absolue) sont observées sur les sites plus arrosés, indiquant une photosynthèse plus élevée dans ces conditions, tandis que les plus faibles valeurs de la GPP (en valeur absolue) concernent les sites moins arrosés. Ceci suggère qu’en Afrique de l’Ouest, le contrôle principal de la variabilité spatiale de la GPP est assuré par les précipitations et la disponibilité des ressources en eau (Hanan et al., 1998 ; Brümmer et al., 2008 ; Williams et al., 2008 ; Ciais et al., 2009 ; Weber et al., 2009 ; Jung et al., 2011 ; Sjöström et al., 2013 ; Ago et al., 2014 ; Ago et al., 2015 ; Quansah et al., 2015), comme l’ont également mis en évidence d’autres auteurs ailleurs dans le monde (Law et al., 2002 ; Luyssaert et al., 2007 ; Archibald et al., 2009). Ceci est parfaitement en accord avec le fait que les MAP déterminent en un endroit donné non seulement le type et la densité de végétation, mais également la structure de la canopée (Aubréville, 1949 ; Weber et al., 2009 ; Kaptue et al., 2010), ce qui est confirmé par la corrélation significative positive (R2 = 0,30 et p = 0,02) trouvée entre le LAImax et la MAP des sites considérés (Figure 6). Cependant, cette corrélation entre la végétation et la MAP sur le long terme tend à se dégrader dans le temps à cause des pressions anthropiques sans cesse croissantes (Hountondji et al., 2006 ; Abdi et al., 2014). Dans la région soudano-sahélienne, une large variabilité inter-annuelle de la GPP corrélée plus avec la MAP que la MAT a également été trouvée par d’autres auteurs (Williams et al., 2008 ; Ciais et al., 2009 ; Weber et al., 2009 ; Jung et al., 2011). Les sites recevant plus de 900 mm de précipitation par an montrent une grande variabilité de la GPP avec des valeurs se situant entre -27,0 (Bellefoungou) et -6,3 t C·ha-1·an-1 (Doguè1). En particulier, plusieurs parmi ces sites recevant une MAP identique (Doguè 1 et 2, par exemple) ont des valeurs différentes de GPP (Figure 4), suggérant ainsi l’influence d’autres variables comme le type de végétation, le degré de perturbation, le mode de gestion et l’usage des écosystèmes. L’évolution de la GPP sur une fonction exponentielle croissante de la MAP suggère également une saturation de la GPP aux MAP plus élevées. Ceci ne pourrait être vérifié que si des estimations de la GPP étaient disponibles sur des sites de la région guinéenne (MAP supérieure à 1 600 mm). Une saturation de la GPP suivant la MAP a été néanmoins mise en évidence dans le monde par plusieurs auteurs à partir des valeurs de MAP de 1 500, 3 000 et 4 000 mm de (Lieth, 1973 ; Luyssaert et al., 2007 ; Garbulsky et al., 2010).
5. Séquestration annuelle de carbone des écosystèmes ouest-africains
27En Afrique de l’Ouest, très peu d’études ont porté sur les flux de carbone dans les écosystèmes et en particulier sur les séquestrations annuelles (Hanan et al., 1998 ; Brümmer et al., 2008 ; Merbold et al., 2009 ; Ago et al., 2014 ; Quansah et al., 2015 ; Tagesson et al., 2015). Néanmoins, quelques résultats d’estimations ou de mesures dans quelques écosystèmes avec différentes méthodes sont représentés sur la figure 7 et résumés dans le tableau 2. Du fait de la non-disponibilité des incertitudes sur les valeurs estimées de la NPP et les mesures de la Rs trouvées dans la littérature concernant certains sites, nous nous focaliserons dans la discussion de cette partie sur les valeurs de NEE calculées à l’aide des mesures directes des échanges nets effectuées avec la technique d’eddy covariance.
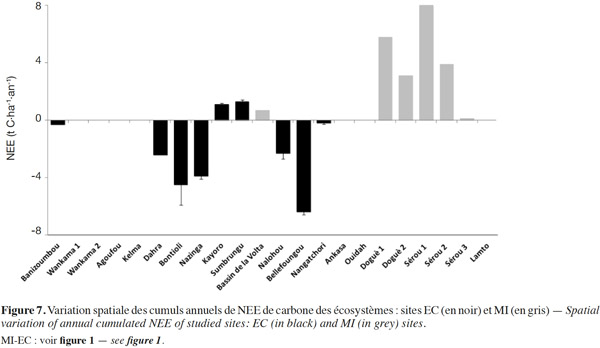
28Les valeurs de NEE estimées à partir des méthodes indirectes sont positives variant entre + 0,1 t C·ha-1·an-1 (superficies agricoles) à Sérou et + 8,0 (forêt dense semi-décidue) au Bénin, suggérant a priori un comportement de source de carbone. Mais l’impossibilité d’estimer les incertitudes sur les valeurs moyennes des composantes majeures des flux nets de carbone (GPP et TER) et donc des NEE annuels de ces sites ne permet pas de tirer une conclusion pertinente. Les deux écosystèmes de forêt à Doguè 1 et Sérou 1 au Bénin avaient été néanmoins observés dans leur phase de vieillissement pendant l’année 2002 par Mulindabigwi (2005), phase essentiellement caractérisée par une décroissance de la GPP ou NPP (en valeur absolue) et une croissance de la TER ou Rh de l’écosystème (DeLucia et al., 2007 ; Amiro et al., 2010).
29L’évolution de la NEE annuelle suivant ses deux principaux composants (GPP, TER) et les facteurs climatiques (MAP, MAT) est présentée à la figure 8. On observe une différenciation de quatre catégories d’écosystèmes en fonction du degré de perturbation et du mode de gestion des écosystèmes à partir des valeurs de la NEE. Ainsi, on quitte les sites protégés, très faiblement ou non perturbés (NEE négatifs plus faibles) vers les sites où la présence d’activités humaines est très marquée (NEE positifs plus élevés). Par ailleurs, aucune dépendance claire de la NEE n’est observée avec la MAT et la MAP (Figure 8b et 8d), confirmant que la variabilité spatiale de la NEE n’est pas principalement déterminée par les facteurs climatiques, mais beaucoup plus par d’autres facteurs non climatiques comme le type de sol, le type de végétation, la structure de la canopée, la profondeur de la nappe, le degré de perturbation, la forme d’utilisation et de gestion des écosystèmes (Hanan et al., 1998 ; Valentini et al., 2000 ; Law et al., 2002 ; Luyssaert et al., 2007 ; Archibald et al., 2009 ; Bombelli et al., 2009 ; Ciais et al., 2011 ; Jung et al., 2011 ; Grace et al., 2014 ; Mbow, 2014 ; Valentini et al., 2014). En effet, les sites de Bellefoungou, de Bontioli et de Nazinga (réserves protégées) se sont comportés comme des puits de carbone, ceux de Dahra et de Nalohou faiblement perturbés (intensité relativement faible des activités humaines) sont des puits modérés de carbone, ceux de Banizoumbou, de Nangatchori moyennement perturbés sont en équilibre et enfin ceux de Sumbrungu et de Kayoro très perturbés (forte intensité des pâtures, feux et activités agricoles) sont de nettes sources de carbone. Toutefois, la moyenne annuelle de NEE, en considérant l’ensemble des sites, est -2,0 ± 1,5 t C·ha-1, ce qui confirme le comportement global de puits de carbone des écosystèmes ouest-africains en parfait accord avec les résultats reportés à l’échelle régionale par d’autres auteurs à partir d’approches de modélisation (Luyssaert et al., 2007 ; Bombelli et al., 2009 ; Weber et al., 2009 ; Jung et al., 2011 ; Fisher et al., 2013). Cependant, il est légitime de craindre à l’avenir que cette région de l’Afrique ne devienne une source de carbone, compte tenu de l’intensité des pressions humaines sans cesse croissantes sur les écosystèmes (Mulindabigwi, 2005 ; Saxton et al., 2007 ; Wittig et al., 2007 ; Bombelli et al., 2009 ; Ozer et al., 2010 ; Ciais et al., 2011 ; Houghton et al., 2012 ; GIEC, 2013 ; Adbi et al., 2014 ; Grace et al., 2014 ; Mbow, 2014 ; Valentini et al., 2014).
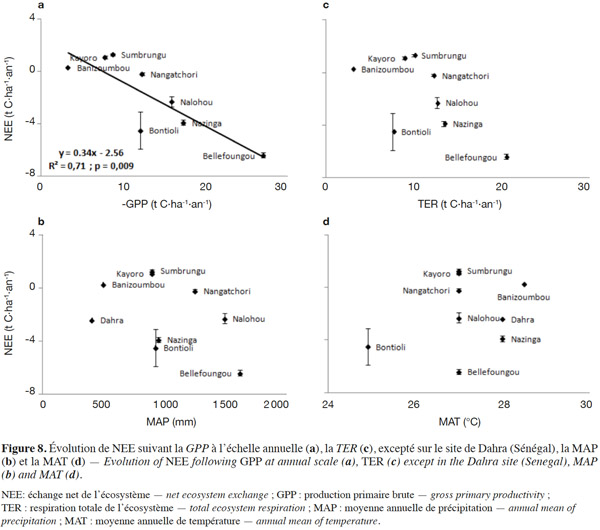
30Par ailleurs, pendant qu’une corrélation significative (R2 = 0,71 ; p = 0,009) a été trouvée entre la NEE et la GPP, une corrélation non significative (R2 = 0,39 ; p = 0,10) a été observée entre la NEE et la TER. Ceci suggère que la variabilité spatiale de la NEE des écosystèmes terrestres ouest-africains est principalement déterminée par celle de la GPP, comme observée dans d’autres régions en Afrique par d’autres auteurs (Weber et al., 2009 ; Jung et al., 2011) et dans le monde (Law et al., 2002 ; Luyssaert et al., 2007). La variabilité spatiale de la GPP apparait plus importante que celle de la TER (Figures 3 et 4), probablement à cause de la forte dépendance de la photosynthèse aux conditions climatiques notamment MAP dans cette région, en comparaison à la TER qui apparait plutôt limitée.
6. Conclusions et perspectives
31Les différents programmes mis en œuvre depuis les années 1970 en Afrique de l’Ouest ont amélioré la compréhension de la dynamique spatio-temporelle des échanges de carbone entre les écosystèmes terrestres et l’atmosphère, et des processus ou facteurs responsables.
32En Afrique de l’Ouest, les moyennes annuelles de la GPP et de la TER sont dépendantes des précipitations moyennes annuelles. En revanche, aucune relation claire n’a été trouvée avec la moyenne annuelle de la température. De même, aucune relation claire n’a été trouvée entre la GPP et la radiation globale des sites. D’autres facteurs non climatiques tels que le type de végétation, le degré de perturbation, le type de sols, la structure de la canopée, la profondeur de la nappe, le mode de gestion et la forme d’utilisation des différents écosystèmes influencent également la variabilité spatiale de la GPP et de la TER.
33Globalement, les écosystèmes ouest-africains étudiés se comportent jusqu’à présent comme des puits de carbone. Les cumuls annuels de flux net de carbone (NEE) apparaissent déterminés par la variabilité de la productivité des écosystèmes (GPP), tandis qu’aucune corrélation significative n’a été trouvée avec les principaux facteurs climatiques (pluviométrie, température et radiation). Il apparait donc clair que les facteurs non climatiques comme le type de végétation, le type de sol, la profondeur de la nappe, le degré de perturbation, la structure de la canopée, l’usage et le mode de gestion des écosystèmes devront être intégrés dans les prochaines études de flux de carbone et des facteurs contrôlant la dynamique de ceux-ci en Afrique de l’Ouest. Ainsi, le contrôle des principaux facteurs responsables des émissions de carbone, en particulier les fortes pressions anthropiques (changements d’utilisation des sols, déforestation, feu, agriculture, pâturage, etc.), sera déterminant pour l’adaptation de ces sites aux changements climatiques et aux évènements météorologiques extrêmes.
34Cependant, il reste bien des aspects nécessitant des études et des recherches plus approfondies. Au delà de l’amélioration de la compréhension des échanges de carbone dans les écosystèmes, l’amélioration des modèles de prédiction des évolutions des flux dans le contexte des variabilités climatiques constituera également l’un des points importants à aborder. Aussi, est-il pertinent d’intensifier les recherches et les mesures des flux de CO2 dans tous les types d’écosystèmes représentatifs de l’Afrique de l’Ouest. Ceci permettra de mieux clarifier les causes des variabilités spatio-temporelles, les différents facteurs pédo-climatiques et les principaux mécanismes responsables. Enfin, la contribution au cycle global de carbone de cette région pourra être évaluée.
35Abréviations
36GPP : production primaire brute
37LAI : index de la surface foliaire
38MAP : moyenne annuelle de précipitation
39MARg : moyenne annuelle de radiation globale
40MAT : moyenne annuelle de température
41NEE : échange net de l’écosystème
42NPP : production primaire nette
43Ra : respiration autotrophe
44Rh : respiration hétérotrophe
45Rs : respiration du sol
46TER : respiration totale de l’écosystème
47Remerciements
48Cette étude a été financée par l’Agence Universitaire de la Francophonie (AUF) et l’Association pour l’Agriculture et le Développement Durable (A2D). Enfin, le premier auteur est reconnaissant envers Ir. Félicien Tosso pour son appui à la réalisation de la carte de localisation des sites et Dr. Ir. Kasso Daïnou pour ses divers conseils et appuis.
Bibliographie
Abdi A.M. et al., 2014. The supply and demand of net primary production in the Sahel. Environ. Res. Lett., 9, 094003.
Ago E.E., Petit F. & Ozer P., 2005. Analyse des inondations en aval du barrage hydroélectrique de Nangbeto sur le fleuve Mono (Togo et Bénin). Geo-Eco-Trop, 29, 1-14.
Ago E.E. et al., 2014. Long term observations of carbon dioxide exchange over cultivated savanna under a Sudanian climate in Benin (West Africa). Agric. For. Meteorol., 197, 13-25.
Ago E.E. et al., 2015. Carbon dioxide fluxes from a degraded woodland in West Africa and their responses to main environmental factors. Carbon Balance Manage., 10, 1-16.
Ajonina G.N. et al., 2014. Carbon budget as a tool for assessing mangrove forests degradation in the western, coastal wetlands complex (Ramsar Site 1017) of Southern Benin, West Africa. In: Diop S., Barusseau J.-P. & Descamps C., eds. The land/ocean interactions in the coastal zone of west and central Africa. Estuaries of the world. Switzerland: Springer International Publishing.
Amiro B.D. et al., 2010. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res., 115, G00K02.
Archibald S.A. et al., 2009. Drivers of inter-annual variability in Net Ecosystem Exchange in a semi-arid savanna ecosystem, South Africa. Biogeosciences, 6, 251-266.
Aubinet M., Vesala T. & Papale D., 2012. Eddy covariance: a practical guide to measurement and data analysis. Dordrecht, The Netherlands: Springer.
Aubréville A., 1949. Climats, forêts et désertification de l’Afrique tropicale. Paris : Société d’éditions géographiques, maritimes et coloniales.
Bombelli A. et al., 2009. An outlook on the sub-Saharan Africa carbon balance. Biogeosciences, 6, 2193-2205.
Bond-Lamberty B., Wang C. & Gower S.T., 2004. A global relationship between the heterotrophic and autotrophic components of soil respiration. Global Change Biol., 10, 1756-1766.
Brümmer C. et al., 2008. Diurnal, seasonal and interannual variation in carbon dioxide and energy exchange in shrub savanna in Burkina Faso (West Africa). J. Geophys. Res., 113, G02030.
Canadell J.G., Raupach M.R. & Houghton R.A., 2009. Anthropogenic CO2 emissions in Africa. Biogeosciences, 6, 463-468.
Carvalho J.A. Jr et al., 1998. Combustion completeness in a rainforest clearing experiment in Manaus, Brazil. J. Geophys. Res., 103(D11), 13195-13199.
Chambers J.Q. et al., 2004. Respiration from a tropical forest ecosystem: partitioning of sources and low carbon use efficiency. Ecol. Appl., 14, 572-588.
Ciais P. et al., 2009. Variability and recent trends in the African carbon balance. Biogeosciences, 5, 3497-3532.
Ciais P. et al., 2011. The carbon balance of Africa: synthesis of recent research studies. Philos. Trans. R. Soc. London, Ser. A, 369, 2038-2057.
Curiel Yuste J. et al., 2007. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Global Change Biol., 13, 2018-2035.
Davidson E.A., Janssens I.A. & Luo Y., 2006. On the variability of respiration in terrestrial ecosystem: moving beyond Q10. Global Change Biol., 12, 154-164.
DeLucia E.H., Drake J.E., Thomas R.B. & Conzalez-Meler M., 2007. Forest carbon use efficiency: is respiration a constant fraction of gross primary production? Global Change Biol., 13, 1157-1167.
Fisher J.B. et al., 2013. African tropical rainforest net carbon dioxide fluxes in the twentieth century. Philos. Trans. R. Soc. London, Ser. B, 368, 20120376.
Fluxnet, 2015. Sites of networks, http://fluxnet.ornl.gov/maps-graphics, (02.03.2015).
Freitag H. et al., 2008. Water and carbon fluxes from savanna ecosystems of the Volta River watershed, West Africa. Global Planet. Change, 61, 3-14.
Friborg T., Boegh E. & Soegaard H., 1997. Carbon dioxide flux, transpiration and light response of millet in the Sahel. J. Hydrol., 189, 633-650.
Garbulsky M.F. et al., 2010. Patterns and controls of the variability of radiation use efficiency and primary productivity accross terrestrial ecosystems. Global Ecol. Biogeogr., 19, 253-267.
GIEC, 2013. Changements climatiques 2013 : les éléments scientifiques. Résumé à l’intention des décideurs. Genève, Suisse : GIEC.
Gonzalez P., Tucker C. & Sy H., 2012. Tree density and species decline in the African Sahel attributable to climate. J. Arid Environ., 78, 55-64.
Grace J., Mitchard E. & Gloor E., 2014. Perturbations in the carbon budget of the tropics. Global Change Biol., 20, 3238-3255.
Hanan N.P., Kabat P., Dolman A.J. & Elbers J.A., 1998. Photosynthesis and carbon balance of a Sahalian fallow savanna. Global Change Biol., 4, 523-538.
Hanson P.J., Edwards N.T., Garten C.T. & Andriew J.A., 2000. Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry, 48, 115-146.
Hiernaux P. et al., 2009. Sahelian rangeland response to changes in rainfall over two decades in the Gourma region, Mali. J. Hydrol., 375, 114-127.
Houghton R.A. et al., 2012. Carbon emissions from land use and land-cover change. Biogeosciences, 9, 5125-5142.
Hountondji Y.C., Sokpon N. & Ozer P., 2006. Analysis of the vegetation trends using low resolution sensing data in Burkina Faso (1982-1999) for the monitoring of desertification. Int. J. Remote Sens., 27(5), 871-884.
Jarvis P. et al., 2007. Dry and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the “Birch effect”. Tree Physiol., 27, 929-940.
Jung M. al., 2011. Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J. Geophys. Res., 116, G00J07.
Kaptue T.A.T., Roujean J.-L. & Faroux S., 2010. ECOCLIMAP II: an ecosystem classification and land surface parameters database of Western Africa at 1 km resolution for the African Monsoon Multidisciplinary Analysis (AMMA) project. Remote Sens. Environ., 114, 961-976.
Lamade E., Djegui N. & Leterme P., 1996. Estimation of carbon allocation to the roots from soil respiration measurements of oil palm. Plant Soil, 181, 329-339.
Landsberg J.J. & Waring R.H., 1997. A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. For. Ecol. Manage., 95, 209-228.
Law B.E. et al., 2002. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agric. For. Meteorol., 113, 97-120.
Le Lay M., 2006. Modélisation hydrologique dans un contexte de variabilité hydro-climatique. Une approche comparative pour l’étude du cycle hydrologique à méso-échelle au Bénin. Thèse de doctorat : Institut National Polytechnique de Grenoble (France).
Lieth H., 1973. Primary production: terrestrial ecosystems. Hum. Ecol., 1(4), 303-332.
Liu H.S. et al., 2006. Respiratory substrate availability plays a crucial role in the response of soil respiration to environmental factors. Appl. Soil Ecol., 32, 284-292.
Loescher H.W. et al., 2003. Environmental controls on net ecosystem-level carbon exchange and productivity in a Central American tropical wet forest. Global Change Biol., 9, 396-412.
Luyssaert S. et al., 2007. The CO2-balance of boreal, temperate and tropical forests derived from a global database. Global Change Biol., 13, 2509-2537.
Malhi Y., 2012. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol., 100, 65-75.
Malhi Y., Baldocchi D.D. & Jarvis P.G., 1999. The carbon balance of tropical, temperate and boreal forests. Plant Cell Environ., 22, 715-740.
Mbow C., 2014. Africa’s greenhouse-gas budget is in the red. Nature, 508, 192-193.
Merbold L. et al., 2009. Precipitation as driver of carbon fluxes in 11 African ecosystems. Biogeosciences, 6, 1027-1041.
MillerA. et al., 2005. Episodic rewetting enhances carbon and nitrogen release from chaparral soils. Soil Biol. Biochem., 37, 2195-2204.
Mordelet P. & Menaut J.C., 1995. Influence of trees on aboveground production dynamics of grasses in a humid savanna. J. Veg. Sci., 6, 223-228.
Mulindabigwi V., 2005. Influence des systèmes agraires sur l’utilisation des terroirs, la séquestration du carbone et la sécurité alimentaire dans le bassin versant de l’Ouémé supérieur au Bénin. Thèse de doctorat : Université de Bonn (Allemagne).
Nemani R.R. et al., 2003. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 300, 1560-1563.
Nicholson S.E., 2013. The West African Sahel: a review of recent studies on the rainfall regime and its interannual variability. ISRN Meteorol., 2013, ID 453521.
Ozer P. et al., 2010. Désertification au Sahel : historique et perspectives. BSGLg, 54, 69-84.
Quansah E. et al., 2015. Carbon dioxide fluxes from contrasting ecosystems in the Sudanian savanna in West Africa. Carbon Balance Manage., 10, 1-17.
Reichstein M. et al., 2003. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Global Biogeochem. Cycles, 17(4), 1-15.
Richardson A.D. et al., 2006. Comparing simple respiration models for eddy flux and dynamic chamber data. Agric. For. Meteorol., 141, 219-234.
Saxton J.E. et al., 2007. Isoprene and monoterpene measurements in secondary forest in northern Benin. Atmos. Chem. Phys., 7, 4095-4106.
Schlesinger W.H., 1997. Biogeochemistry: an analysis of global change. New York, USA: Academic Press.
Schulze E.-D., 2006. Biological control of the terrestrial carbon sink. Biogeosciences, 3, 147-166.
Seghieri J. et al., 2009. Relationships between climate, soil moisture and phenology of the woody cover in two sites located along the West African latitudinal gradient. J. Hydrol., 375, 78-89.
Sjöström M. et al., 2013. Evaluation of MODIS gross primary productivity for Africa using eddy covariance data. Remote Sens. Environ., 131, 275-286.
Tagesson T. et al., 2015. Ecosystem properties of semiarid savanna grassland in West Africa and its relationship with environmental variability. Global Change Biol., 21, 250-264.
UNDP, 2015. World population prospects: the 2012 revision, http://esa.un.org/unpp, (26.11.2015).
Valentini R. et al., 2000. Respiration as the main determinant of carbon balance in European forests. Nature, 404, 861-865.
Valentini R. et al., 2014. A full greenhouse gases budget of Africa: synthesis, uncertainties and vulnerabilities. Biogeosciences, 11, 381-407.
Weber U. et al., 2009. The interannual variability of Africa’s ecosystem productivity: a multi-model analysis. Biogeosciences, 6, 285-295.
Williams C.A. et al., 2008. Interannual variability of photosynthesis across Africa and its attribution. J. Geophys. Res., 113, G04015.
Williams C.A. et al., 2009. Complexity in water and carbon dioxide fluxes following rain pulses in an African savanna. Oecologia, 161, 469-480.
Wittig R., König K., Schmidt M. & Szarzynski J., 2007. A study of climate change and anthropogenic impacts in West Africa. Environ. Sci. Pollut. Res., 14(3), 182-189.
