


- Portada
- Volume 19 (2015)
- numéro 4
- Meliponini and Apini in Africa (Apidae: Apinae): a review on the challenges and stakes bound to their diversity and their distribution


Vista(s): 4294 (77 ULiège)
Descargar(s): 464 (10 ULiège)

Meliponini and Apini in Africa (Apidae: Apinae): a review on the challenges and stakes bound to their diversity and their distribution

Notes de la rédaction
Received on August 27, 2014; accepted on April 3, 2015
Résumé
Les Meliponini et les Apini en Afrique (Apidae : Apinae) : synthèse bibliographique sur les défis et les enjeux liés à leur diversité et leur distribution
Introduction. Les Meliponini et les Apini contribuent à la pollinisation des plantes à fleurs et à l’amélioration des rendements agricoles. La diversité, la distribution et l’abondance de ces abeilles dépendent de l’écosystème dans lequel elles évoluent. Le présent travail vise à faire une synthèse sur la diversité, la distribution, l'abondance, les menaces potentielles, les enjeux et les défis rencontrés à l'égard de ces abeilles sociales en Afrique.
Littérature. En Afrique, on a dénombré 21 espèces de Meliponini et 2 espèces d’Apini (Apis). La diversité, la distribution, la biologie, l’écologie et l’abondance de ces espèces sont peu documentées, particulièrement pour les Meliponini. Cela pourrait être lié au manque d’intérêt des auteurs ou aux différentes difficultés pour réaliser ces études en Afrique. En ce qui concerne les difficultés, il est nécessaire de clarifier la taxonomie des Meliponini et l'observation de nids dans certains milieux forestiers est difficile. La prédation des nids et la perte d’habitat restent les principales menaces qui pourraient occasionner le dépeuplement de certaines espèces d’abeilles sociales en Afrique. Si les maladies et les parasites ne constituent pas encore une grande menace pour ces abeilles, il est nécessaire de poursuivre de nouvelles études pour mieux évaluer les dangers liés à ces potentielles menaces.
Conclusions. Les travaux sur la diversité, la distribution et l’abondance des abeilles sociales doivent être renforcés afin de relever les défis qui sont liés à ces insectes en Afrique. En effet, cela contribuera à répondre à des enjeux de gestion durable de la biodiversité et à des enjeux économiques et agricoles.
Abstract
Introduction. Meliponini and Apini contribute to the pollination of flowering plants and to improving agricultural yields. These bees’ diversity, distribution and abundance depend on the ecosystem in which they evolve. The present work aims to summarize the diversity, distribution, abundance, potential threats, challenges and issues faced with respect to these social bees in Africa.
Literature. In Africa, there were 21 species of Meliponini and 2 species of Apini (Apis). Aspects related to the species diversity, distribution, biology, ecology and abundance of nests are poorly documented, especially for Meliponini. This deficit could be related to a lack of interest of the authors or the various difficulties in performing these studies in Africa. With regard to the difficulties, there is a need to clarify the taxonomy of Meliponini, and the observation of nests in certain forest environments is difficult. Nest predation and habitat loss are the main threats that could cause the depopulation of certain social bee species in Africa. If there is currently new evidence that diseases and pests did not endanger these bees, then there is a need for further studies for better assessment of the risks that are associated with these potential threats.
Conclusions. Work on the diversity, distribution and abundance of the social bees must be strengthened to address the challenges that are related to these insects in Africa. Indeed, this approach will contribute to answering the challenges of sustainable management of the biodiversity and economic and agricultural issues.
Tabla de contenidos
1. Introduction
1Bees play a role in the reproduction of some flowering plants, similar to other pollinators (Bradbear, 2010; Ollerton et al., 2011). Through this activity, bees contribute to the preservation and maintenance of the genetic diversity of flowering plants (Bradbear, 2010). In agriculture, bees contribute to an increase in the qualitative and quantitative crop yields (Klein et al., 2007; Klatt et al., 2014). While bees contribute to the preservation of ecosystems, they are also dependent on the balance of these ecosystems. Therefore, their diversity, distribution and abundance are related to different environmental conditions as well as the impact of human activity (Brown et al., 2009). Among these arthropods, two groups can be distinguished according to their social organization: solitary and social. The subject of this paper focuses on Meliponini and Apini, which belong to the social bees group (highly eusocial). These bees are organized as colonies that consist of a female queen, males, and female workers that are responsible for different activities (Michener, 2007). Threats to social bees, which are related to diseases, parasites and habitat loss, have been reported worldwide (Jaffé et al., 2009; Van Der Zee et al., 2012). In Africa, there is less scientific work that has been published on highly eusocial, especially for Meliponini, of which the taxonomy still must be clarified (Eardley et al., 2013). Considering the ecological and economic importance of bees, it is important to gain knowledge about them in Africa. This concern is especially important because this continent contains the second largest forest in the world, in which there is a high level of ecosystem diversity.
2The aim of this study is to summarize the current knowledge on the species diversity, distribution and abundance of Meliponini and Apini in Africa in general and in Gabon, a country in the Congo Basin Forest, in particular. Potential threats to the diversity of social bees will be discussed as well as the environmental and economic challenges and prospects that are related to these insects in Africa.
2. Highly eusocial bees: diversity, distribution, and abundance
2.1. The Meliponini
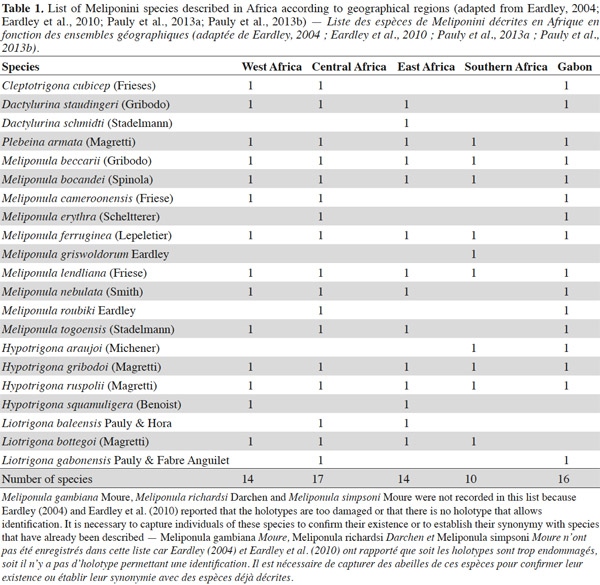
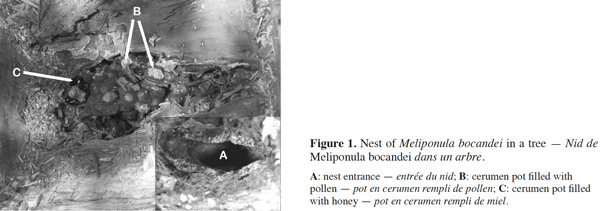
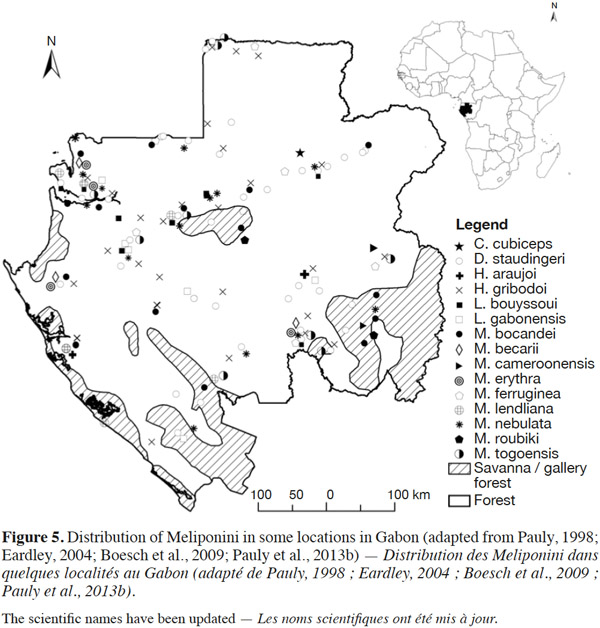
3In Africa, 21 species of Meliponini have been listed (Table 1) and divided into six genera: Cleptotrigona, Dactylurina, Hypotrigona, Liotrigona, Meliponula, and Plebeina (Eardley, 2004; Eardley et al., 2010). Although the Meliponini are present in savannahs, many species nest mainly in forested areas. The forest species, such as Meliponula beccarii (Gribodo) and Meliponula bocandei (Spinola), nest on the ground or in trees (Kajobe, 2007a; Mogho Njoya, 2009) (Figure 1). Other species, such as Meliponula nebulata (Smith), nest in the cavities of trees or in termite mounds (Brosset et al., 1967; Darchen, 1969; Kajobe, 2007a; Mogho Njoya, 2009) (Figures 2 and 3). Central African countries have the highest reported diversity of Meliponini, with 17 species (Table 1); 16 of these 17 species have been reported in Gabon (Table 1) (Figure 4). Of all of the species listed in Gabon, Meliponula roubiki Eardley has been described and is found only in this country in Africa (Eardley, 2004; Eardley et al., 2010) (Table 1). Pauly’s (1998) inventory showed that many Meliponini species in Gabon were distributed throughout the entire territory, including Dactylurina staudingeri (Gribodo), M. bocandei, M. nebulata, and the genus Hypotrigona (Figure 5). Other species have been inventoried in smaller regions (i.e., Meliponula cameroonensis (Friese) and Meliponula lendliana (Friese)) (Figure 5).



4The actual number of species requires revision because the taxonomy of the Meliponini in Africa is unclear, and some cryptic species remain to be identified on the African continent (Eardley et al., 2013). The description of the Hypotrigona and Liotrigona species, in particular, requires clarification.
5Unlike in the case of the Meliponini of South America or Asia, there are few published studies on aspects of morphometrics that relate to the Meliponini in Africa. Darchen (1969; 1971) performed body measurements on Meliponula ferruginea (Lepeletier) (synonymous with Meliponula oyani [Darchen]) (Eardley et al., 2010)) and M. nebulata in Gabon. These measurements were performed without statistical analysis but within the framework of a description of these species; the results showed great variability for M. nebulata. In Ghana, Combey et al. (2013) studied biodiversity in four species of Meliponini (M. ferruginea, M. bocandei, D. staudingeri, and Hypotrigona gribodoi [Magretti]). Using geometric morphometrics, the authors demonstrated the existence of possible ecotypes within these species. The study focused on the use of landmarks that were positioned on the right forewings of these bees.
6Research on the genetic diversity of the Meliponini in Africa is also poorly documented. Several African species have been studied molecularly by Costa et al. (2003) and Rasmussen et al. (2010). Both sets of authors studied phylogenetic species evolution on a global scale, including African species, such as Meliponula ogouensis (Vachal) (synonymous with Meliponula becarri [Gribodo] [Pauly et al., 2013a]), M. bocandei, M. ferruginea, M. lendliana, D. staudingeri, H. gribodoi and Hypotrigona ruspolii (Magretti), and showed that Hypotrigona was distant from other African genera.
7Regarding the abundance of Meliponini, Darchen (1972) reported a nest density of 2.5 nests·ha-1, mostly containing the genera Hypotrigona, Liotrigona and Dactylurina, in Lamto savannahs in Côte d’Ivoire. In Ugandan forests, Kajobe et al. (2006) reported a density of 0.39 nests·ha-1 of highly eusocial bees, of which 0.27 nests·ha-1 contained Meliponini. In addition, Tornyie et al. (2015) found a Meliponini nest density of between 1.7 and 2.4 nests·ha-1 in Ghana. This study focuses on M. bocandei, M. ferruginea and D. staudingeri. The difference between the results of these studies might relate to differences in the habitat studied and the sampling effort employed in the different countries.
8These studies are representative of the sparse data regarding social bees within the 54 countries of Africa. This lack of data could reflect a low level of interest among researchers or the difficulties encountered in studying the diversity and abundance of social bees in Africa. Indeed, the difficulties that are involved in assessing the overall diversity and abundance of these bees in this continent are numerous, and several are common to many African countries. One major difficulty is the uncertain taxonomy of specific Meliponini species, which would not facilitate the study of the diversity, biology, ecology, and other aspects of these insects (Eardley et al., 2013).
9Political instability and civil or communitarian wars in some parts of Africa constitute another difficulty for the study of the diversity and abundance of bees. Access to certain areas is often dangerous; however, Gabon is less affected by this problem because it has a relatively stable political situation.
10Given a high plant density and rugged terrain, the access to certain forested areas in Gabon is difficult. In addition, the canopy height makes it difficult to study these insects (Pauly, 1998). A large number of bee colonies reside in tall trees and are difficult to reach. Furthermore, the bees forage on flowers that are blooming in the canopy, which makes it difficult to observe or capture these insects.
2.2. The Apini
11In Africa, two species have been listed within the Apis genus: Apis florea Fabricius and Apis mellifera Linnaeus (Eardley et al., 2010; Hepburn et al., 2011; Pauly et al., 2013a). Apis florea is distributed from Asia to east Africa (Hepburn et al., 2011), while A. mellifera is subdivided into 10 identified subspecies: A. mellifera adansonii Latreille, A. mellifera capensis Eschscholtz, A. mellifera intermissa Von Buttel-Reepen, A. mellifera jemenitica Ruttner, A. mellifera lamarckii Cockerell, A. mellifera litorea Smith, A. mellifera monticola Smith, A. mellifera sahariensis Baldensperger, A. mellifera scutellata Lepeletier and A. mellifera simensis Meixner et al. (Hepburn et al., 1998; Meixner et al., 2011). These subspecies have been described from morphological and traditional morphometric studies (Smith, 1961; Ruttner, 1988; Meixner et al., 2011). Ruttner (1988) determined that the races or subspecies of honeybee can be classified into four main evolutionary lineages: M, C, O, and A. African honeybees belong to a single evolutionary line (A) as mentioned by Ruttner (1988) and confirmed by the geometric morphometric studies of Kandemir et al. (2011).
12Molecular analysis and nest abundance of honeybees in Africa have not yet been well documented. Franck et al. (2001) and Jaffé et al. (2009) showed that honeybees presented greater genetic diversity in Africa than in Europe. Gruber et al. (2013) suggested that the morphometric differences between honeybees in the disjunct montane forests (A. mellifera monticola) and those in the savannahs (A. mellifera scutellata) in Kenya were more closely related to a phenomenon of phenotypic plasticity. The molecular analyses that were performed by these authors showed no distinction between these honeybees in both of these locations, which allowed these bees to be divided into two subspecies, as suggested by morphometric analysis. If these results are confirmed by other studies, the existence of A. mellifera monticola, a honeybee that is present in mountain refugia, could be revisited.
13Regarding the nest density, in Botswana, McNally et al. (1996) found a density of approximately 0.04 nest·ha-1. In Uganda, Kajobe et al. (2006) found a nest abundance of 0.12 nest·ha-1 for A. mellifera. Jaffé et al. (2009) used the distance covered by the drones and the queens to participate in the coupling process to estimate the density of honeybee colonies in various localities in Africa, Europe and Asia. The nest density ranged from 0.052 to 0.097 nest·ha-1 in the localities studied in South Africa; the density was 0.042 nest·ha-1 in Sudan and 0.062 nest·ha-1 in Libya (Jaffé et al., 2009). While the taxonomy might not be a problem here as it is with the Meliponini, problems that are related to political instability in certain regions and the constraints related to the forest environment are major difficulties for the study of the diversity and abundance of honeybees in Africa.
14Only A. mellifera adansonii has been identified in Gabon (Ambougou Atisso, 1991; Pauly, 1998). Traditional morphometric studies of honeybees in several localities in Gabon have shown that the measurements of A. mellifera adansonii are homogeneous across the country (Radloff et al., 1999). The use of geometric morphometrics to confirm the homogeneity of this subspecies in Gabon could be informative. Unlike some Meliponini species, A. mellifera adansonii has a wide range of habitats in its geographical area, and it builds its nests in a range of cavity types. To date (February 2015), no scientific study has yet been published in Gabon on the molecular diversity or nest abundance of honeybees.
3. Threats to the diversity and abundance of highly eusocial bees
15In Africa, threats to highly eusocial bees largely relate to nest predation and habitat loss (Dietemann et al., 2009); their predators include humans and chimpanzees (Kajobe et al., 2006; McLennan et al., 2011). Humans destroy or disrupt bee colonies when harvesting honey, and the harvesting methods employed can contribute to the depopulation of certain bee species (Bradbear, 2010). However, data do not currently exist regarding the loss of bee colonies as a result of the predation of nests by humans. In certain countries of Africa, the development of beekeeping has helped to improve the incomes of rural populations, but it has also contributed to a reduction in human nest predation (Dietemann et al., 2009; Bradbear, 2010). In Gabon, apiculture is rarely practiced, and the breeding of stingless bees does not take place. Honey hunting mainly occurs in rural areas and is accompanied by the destruction of nests. The only known bee conservation measure in Gabon is the ban on the felling of trees for the purposes of harvesting honey in national parks.
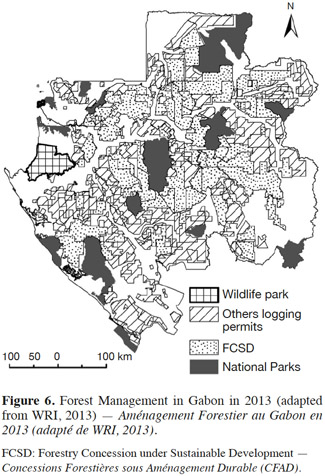
16The loss of habitat for bees in Africa is primarily the result of human activity, due mainly to agriculture and deforestation (Dietemann et al., 2009). In Gabon, forest represents approximately 78% of its territory (Drouineau et al., 1999). Logging concessions cover approximately 51% of the territory, and protected areas (all national parks and Wildlife Parks) cover approximately 13% (adapted from WRI’s data, 2013) (Figure 6). A large number of logging permits is issued by the Forestry Concession under Sustainable Development (FCSD). Gabonese legislation requires that logging companies that hold FCSD permits carry out a management and operating plan of their forest concession to ensure the sustainable management of forest resources and the preservation of biodiversity (Présidence de la République Gabonaise, 2001). Some loggers in Gabon have Forest Stewardship Council (FSC) certifications, which guarantee sustainable forest management. However, the application of these management regimes in Gabon does not take bees into consideration. Indeed, current conservation measures concern only plant and animal species that are completely protected by law or concern delicate ecosystems such as wetlands. It is therefore important to study the influence of these and other human activities on the habitats and diversity of social bees. Many forest trees are pollinated by bees, and if the bees are endangered by logging or other activities, the pollination of these forest plants is also affected.
17Although it has not yet been established that pests and diseases pose a major threat to the diversity of social bees in Africa, it is necessary to continue research into entomopathogens and parasites of both stingless bees and honeybees on the continent. Reports of the United States and Europe being affected by the loss of honeybee colonies underline the importance of this research (Pettis et al., 2010; Van Der Zee et al., 2012). Such research would also provide much needed data for a large number of African countries, where data are currently lacking (Mumoki et al., 2014). Some studies have shown the presence of certain parasites and diseases, such as the invasive varroa mite in Africa (Dietemann et al., 2009). However, it appears that these pests and diseases bear little responsibility for honeybee colony loss in Africa, as demonstrated by the results of Strauss et al. (2013) in South Africa and by Muli et al. (2014) in Kenya. Ellis (2005) showed that honeybees were able to contain the endemic small hive beetle in part of a hive. Mumbi et al. (2014) observed the presence of varroa in many localities in Tanzania. The number of mites per colony count was low compared to those observed in Europe or America, and the colonies showed no visible clinical signs (Mumbi et al., 2014). However, Akinwande et al. (2013) concluded that insect pests, diseases, and pesticide poisoning might be the main factors that underlie the declines in the establishment of honeybee colonies in Nigeria.
18To date, no known studies have been published in Gabon on the pests, diseases, or any other potential threat to stingless bees or honeybees.
4. Diversity, distribution and abundance of highly eusocial bees: what are the challenges and issues that are related to the question of conservation, agriculture and economics?
19The first challenge that is involved in this area of study is to improve the knowledge and the extent of the conservation of the diversity of social bees. This goal is essential and would be beneficial to the maintenance of bee populations but also to the maintenance of the balance within ecosystems due to the role played by bees in pollinating wild plants (Dietemann et al., 2009). Indeed, Fontaine et al. (2006) showed that the loss of the functional diversity of pollinators could cause a loss or decline in the diversity of plants and could consequently affect the composition of the natural plant communities. Unlike some pollinators, social bees visit a variety of plants. Thus, in Uganda, A.mellifera, M. bocandei and M. nebulata visited, respectively, 71, 52 and 33 plants species according to Kajobe (2007b). In Gabon, Ambougou Atisso (1991) found pollen from 151 different plant species in the food reserves collected by A. mellifera adansonii, with 136 taxa represented in honey and 91 in pollen reserves. These plant taxa originated from various habitats, such as plantations, ruderal areas, fallow areas, secondary forests, dense forests, and riparian forests (Ambougou Atisso, 1991). Given these results, it can be observed that preserving ecosystem diversity in Gabon requires both the consideration of bees in conservation strategies and the sustainable management of forest resources. Therefore, an improvement in the availability of information in Gabon regarding bee diversity, ecology, biology, and potential threats is important.
20On agronomic and economic levels, bees improve the quantity and quality of many food and cash-crop yields (Klein et al., 2007; Vanbergen, 2014), such as citrus, coffee, maize, cassava, peanuts, safou, avocado, and many others (Ambougou Atisso, 1991; Tchuenguem Fohouo et al., 2001; Tchuenguem Fohouo et al., 2002), via their beneficial effect on pollination. The combination of crop cultivations and beekeeping could provide an opportunity for Gabon to develop its agriculture and could contribute to improved yields and the diversification of farmers’ incomes.
21Apiculture and stingless beekeeping vary from one country to another; apiculture has been developed in some African countries, such as Uganda, Ethiopia, South Africa, Kenya, and Cameroon (Dietemann et al., 2009), but in Gabon, the practice is still in its infancy. Stingless beekeeping is relatively developed in some countries in South America, such as Brazil (Cortopassi-Laurino et al., 2006). In Africa, stingless beekeeping trials were conducted in Angola, and there is more interest in this breeding in other countries such as Ghana, Kenya, Botswana, and South Africa (Cortopassi-Laurino et al., 2006).
22In Gabon, stingless beekeeping is non-existent, and its development there remains a challenge. Generally, apiculture and stingless beekeeping are based on the use of bait hives or the transfer of wild colonies into hives in Africa (Dietemann et al., 2009). Thus, better knowledge of the diversity, distribution, and abundance of honeybees and stingless bees according to the habitat or localities is essential for an extension of this breeding in Gabon.
23Honey produced by stingless bees is used in traditional medicine by rural African populations. For example, Ewnetu et al. (2013) and Kwapong et al. (2013) showed that honey produced by stingless bees exhibits a superior inhibitory action on certain pathogenic agents (e.g., Staphylococcus, Escherichia, and Klebsiella) in comparison with those of some standard antibiotics. In Gabon, the honey produced by stingless bees and used in traditional medicine is known by two names: sweet honey, for honey produced by M. bocandei, and sour honey, for honey produced by all other stingless bees. Confusion could therefore arise when distinguishing the honey of other species and the medicinal properties of the product. Clarification of the taxonomy and study of the medicinal properties of the honey produced by stingless bees would increase its value.
24Despite the difficulties encountered in the study of African stingless bees and honeybees, research initiatives are currently being undertaken. One example can be observed in the African Pollinator Initiative, which brings together 15 African countries and focuses on the preservation, conservation, and improvement of knowledge regarding pollinators (Byrne et al., 2009). In Gabon, further efforts are required to revive interest in research on social bees, especially because most studies on these insects were conducted more than 30 years ago.
5. Conclusions
25Highly social bees (Meliponini and Apini) have been less studied in Africa. Although the bees’ morphological description has allowed the identification of many Meliponini species, the taxonomy of stingless bees remains to be clarified. As a complementary tool, molecular analysis would be useful for clarifying the identification of stingless bee species in Africa. Morphometry is another tool that would allow the study of the shape and size of the bees according to the species, locality, and ecosystem. Clarification of the taxonomy would facilitate the study of the diversity, biology, and ecology of social bees in Africa.
26The study of potential threats to the Meliponini and Apini must be strengthened in Africa, especially considering the fact that habitat degradation endangers the diversity of social bees on the continent. Pests and diseases represent potential threats, and their impacts must be assessed and monitored in many African countries, including Gabon, where no such data currently exist. In addition, the impact of nest predation on the diversity of social bees during honey hunting also must be evaluated to obtain reliable data on the impact of this activity on social bees in Africa. Alternatives such as the extension of the breeding of social bees, especially in Gabon, should be implemented to reduce the impact of nest predation.
27All of these aspects to be cleared up constitute challenges in relation to environmental issues, with the problems being the conservation of ecosystems, agricultural and economic aspects, better management and better exploitation of the social bees in Africa. The efforts that have already been made in the framework of the study of diversity, distribution and abundance will have to be continued and improved in Africa and especially in Gabon.
28Acknowledgements
29The authors thank the Gabonese government for financially supporting the Ph.D. of Edgard Fabre Anguilet. We also thank the Institut de Pharmacie et de Médecine traditionnelles, Institut de Recherche en Écologie Tropicale, the National Agency of the National Parks, Centre d’Introduction, d’Adaptation et de Multiplication du Matériel Végétal, and Precious Woods Gabon, for their logistical support. Lastly, we also thank Alain Pauly, family and friends for their helpful advice and kind cooperation.
Bibliographie
Akinwande K.L., Badejo M.A. & Ogbogu S.S., 2013. Challenges associated with the honey bee (Apis mellifera adansonii) colonies establishment in south western Nigeria. Afr. J. Food Agric. Nutr. Dev., 13(2), 7467-7484.
Ambougou Atisso V., 1991. Apis mellifera adansonii Lat. et les plantes utiles mellifères gabonaises (Département de l’Ivindo), recherches palynologiques. Thèse de doctorat : Université Paris 6 (France).
Boesch C., Head J. & Robbins M.M., 2009. Complex tool sets for honey extraction among chimpanzees in Loango National Park, Gabon. J. Hum. Evol., 56(6), 560-569.
Bradbear N., 2010. Le rôle des abeilles dans le développement rural. Rome : FAO, http://www.fao.org/docrep/013/i0842f/i0842f00.pdf, (12/11/2011).
Brosset A. & Darchen R., 1967. Une curieuse succession d'hôtes parasites des nids de Nasutitermes. Biol. Gab., 3(2), 153-168.
Brown M.J.F. & Paxton R.J., 2009. The conservation of bees: a global perspective. Apidologie, 40(3), 410-416.
Byrne A. & Fitzpatrick Ú., 2009. Bee conservation policy at the global, regional and national levels. Apidologie, 40(3), 194-210.
Combey R. et al., 2013. Geometric morphometrics reveals morphological differentiation within four African stingless bee species Rofela. Ann. Biol. Res., 4(12), 95-105.
Cortopassi-Laurino M. et al., 2006. Global meliponiculture: challenges and opportunities. Apidologie, 37(2), 275-292.
Costa M.A., Del Lama M.A., Melo G.A.R. & Sheppard W.S., 2003. Molecular phylogeny of the stingless bees (Apidae, Apinae, Meliponini) inferred from mitochondrial 16S rDNA sequences. Apidologie, 34(1), 73-84.
Darchen R., 1969. Sur la biologie de Trigona (Apotrigona) nebulata Komiensis Cock. Biol. Gabonica, 3, 151-183.
Darchen R., 1971. Trigona (Axestotrigona) oyani Darchen (Apidae, Trigoninae), une nouvelle espèce d'abeille africaine - description du nid inclus dans une fourmilière. Biol. Gabonica, 7, 407-421.
Darchen R., 1972. Écologie de quelques trigones (Trigona sp.) de la savane de Lamto (Côte d’Ivoire). Apidologie, 3(4), 341-367.
Dietemann V., Pirk C.W.W. & Crewe R., 2009. Is there a need for conservation of honeybees in Africa? Apidologie, 40(3), 285-295.
Drouineau S. & Robert N., 1999. L’aménagement forestier au Gabon : historique, bilan, perspectives, http://forafri.cirad.fr/ressources/forafri/08.pdf, (10/8/2015).
Eardley C.D., 2004. Taxonomic revision of the African stingless bees (Apoidea: Apidae: Apinae: Meliponini). Afr. Plant Prot., 10(2), 63-96.
Eardley C.D. & Urban R., 2010. Catalogue of Afrotropical bees (Hymenoptera: Apoidea: Apiformes). Zootaxa, 2455, 1-548.
Eardley C.D. & Kwapong P., 2013. Taxonomy as a tool for conservation of African stingless bees and their honey. In: Vita P., Pedro S. & Roubik D.W., eds. Pot-honey: a legacy of stingless bees. New York, NY, USA: Springer, 261-268.
Ellis J.D., 2005. Reviewing the confinement of small hive beetles (Aethina tumida) by western honey bees (Apis mellifera). Bee World, 86(3), 56-62.
Ewnetu Y., Lemma W. & Birhane N., 2013. Antibacterial effects of Apis mellifera and stingless bees honeys on susceptible and resistant strains of Escherichia coli, Staphylococcus aureus and Klebsiella pneumoniae in Gondar, Northwest Ethiopia. BMC Complementary Altern. Med., 13, 269.
Fontaine C., Dajoz I., Meriguet J. & Loreau M., 2006. Functional diversity of plant–pollinator interaction webs enhances the persistence of plant communities. PLoS Biol., 4(1), e1.
Franck P. et al., 2001. Genetic diversity of the honeybee in Africa: microsatellite and mitochondrial data. Heredity, 86(4), 420-430.
Gruber K. et al., 2013. Distinct subspecies or phenotypic plasticity? Genetic and morphological differentiation of mountain honey bees in East Africa. Ecol. Evol., 3(10), 3204-3218.
Hepburn H.R. & Radloff S.E., 1998. Honeybees of Africa. Berlin: Springer Verlag.
Hepburn H.R. & Radloff S.E., 2011. Biogeography of the dwarf honeybees, Apis andreniformis and Apis florea. Apidologie, 42(3), 293-300.
Jaffé R. et al., 2009. Estimating the density of honeybee colonies across their natural range to fill the gap in pollinator decline censuses. Conserv. Biol., 24(2), 583-593.
Kajobe R., 2007a. Nesting biology of equatorial afrotropical stingless bees (Apidae; Meliponini) in Bwindi Impenetrable National Park, Uganda. J. Apic. Res., 46(4), 245-255.
Kajobe R., 2007b. Pollen foraging by Apis mellifera and stingless bees Meliponula bocandei and Meliponula nebulata in Bwindi Impenetrable National Park, Uganda. Afr. J. Ecol., 45(3), 265-274.
Kajobe R. & Roubik D.W., 2006. Honey-making bee colony abundance and predation by apes and humans in a Uganda Forest Reserve. Biotropica, 38(2), 210-218.
Kandemir İ., Özkan A. & Fuchs S., 2011. Reevaluation of honeybee (Apis mellifera) microtaxonomy: a geometric morphometric approach. Apidologie, 42(5), 618-627.
Klatt B.K. et al., 2014. Bee pollination improves crop quality, shelf life and commercial value. Proc. R. Soc. B., 281, 20132440.
Klein A.M. et al., 2007. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B, 274, 303-313.
Kwapong P.K., Ilechie A.A. & Kusi R., 2013. Comparative antibacterial activity of stingless bee honey and standard antibiotics against common eye pathogens. J. Microbiol. Biotechnol. Res., 3(1), 162-168.
Mc Nally L.C. & Schneider S.S., 1996. Spatial distribution and nesting biology of colonies of the African honey bee Apis mellifera scutellata (Hymenoptera: Apidae) in Botswana, Africa. Environ. Entomol., 25(3), 643-652.
McLennan M.R., 2011. Tool-use to obtain honey by chimpanzees at Bulindi: new record from Uganda. Primates, 52(4), 315-322.
Meixner D.M., Leta M.A., Koeniger N. & Fuchs S., 2011. The honey bees of Ethiopia represent a new subspecies of Apis mellifera—Apis mellifera simensis n. ssp. Apidologie, 42(3), 425-437.
Michener C.D., 2007. The bees of the world. 2nd ed. Baltimore, MD, USA: The Johns Hopkins University Press.
Mogho Njoya M.T., 2009. Diversity of stingless bees in Bamenda Afromontane Forests – Cameroon: nest architecture, behaviour and labour calendar. PhD thesis: Wilhelms Universität Bonn - Institut für Nutzpflanzenwissenschaften und Ressourcenschutz Rheinische Friedrich (Deutschland).
Muli E. et al., 2014. Evaluation of the distribution and impacts of parasites, pathogens, and pesticides on honeybee (Apis mellifera) populations in East Africa. PLoS ONE, 9(4), e94459.
Mumbi C.T. et al., 2014. Parasitic mite, Varroa species (Parasitiformes: Varroidae) infesting the colonies of African honeybees, Apis mellifera scutellata (Hymenoptera: Apididae) in Tanzania. J. Entomol. Zool. Stud., 2(3), 188-196.
Mumoki F.N. et al., 2014. An inventory of documented diseases of African honeybees: review article. Afr. Entomol., 22(3), 473-487.
Ollerton J., Winfree R. & Tarrant S., 2011. How many flowering plants are pollinated by animals? Oikos, 120(321), 321-326.
Pauly A., 1998. Hymenoptera Apoidea du Gabon. Ann. Musée R. Afr. Centrale (Sci. Zool.) Tervuren (Belgique), 282, 1-121.
Pauly A. & Hora Z.A., 2013a. Apini and Meliponini from Ethiopia (Hymenoptera: Apoidea: Apidae: Apinae). Belg. J. Entomol., 16, 1-36.
Pauly A. & Fabre Anguilet E.C., 2013b. Description de Liotrigona gabonensis sp. nov., et quelques corrections à la synonymie des espèces africaines de mélipones (Hymenoptera : Apoidea : Apinae : Meliponini). Belg. J. Entomol., 15, 1-13.
Pauly A. & Vereecken N., 2013c. Les Meliponinae africaines, http://www.atlashymenoptera.net/page.asp?ID=121, (03/12/2014).
Pettis J.S. & Delaplane K.S., 2010. Coordinated responses to honey bee decline in the USA. Apidologie, 41(3), 256-263.
Présidence de la République Gabonaise, 2001. Loi n°16/01 portant code forestier en république gabonaise.
Radloff S.E. & Hepburn H.R., 1999. Honeybees, Apis mellifera Linnaeus (Hymenoptera: Apidae), of equatorial Africa. Afr. Entomol., 7(2), 265-269.
Rasmussen C. & Cameron S.A., 2010. Global stingless bee phylogeny supports ancient divergence, vicariance, and long distance dispersal. Biol. J. Linn. Soc., 99(1), 206-232.
Ruttner F., 1988. Biogeography and taxonomy of honeybees. Berlin: Springer-Verlag.
Smith F.G., 1961. The races of honeybees in Africa. Bee World, 42, 255-260.
Strauss U. et al., 2013. Seasonal prevalence of pathogens and parasites in the savannah honeybee (Apis mellifera scutellata). J. Invertebr. Pathol., 114, 45-52.
Tornyie F. & Kwapong P.K., 2015. Nesting ecology of stingless bees and potential threats to their survival within selected landscapes in the northern Volta region of Ghana. Afr. J. Ecol., doi: 10.1111/aje.12208.
Tchuenguem F.-N., Messi J. & Pauly A., 2001. Activité de Meliponula erythra sur les fleurs de Dacryodes edulis et son impact sur la fructification. Fruits, 56(3), 179-188.
Tchuenguem F.-N., Messi J. & Pauly A., 2002. Recherches sur l’activité des Apoïdes sauvages sur le maïs à Yaoundé (Cameroun) et réflexions sur la pollinisation des graminées. Biotechnol. Agron. Soc. Environ., 6(2), 87-98.
Van Der Zee R. et al., 2012. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008-9 and 2009-10. J. Apic. Res., 51(1), 100-114.
Vanbergen A.J., 2014. Landscape alteration and habitat modification: impacts on plant – pollinator systems. Curr. Opin. Insect Sci., 5, 44-49.
WRI (World Resources Institute), 2013. The interactive forest atlas of Gabon, http://www.wri.org/our-work/project/congo-basin-forests/gabon#project-tabs, (16/06/2014).
