


- Accueil
- Volume 18 (2014)
- Numéro 1
- Apport du traitement d’images dans le suivi de l’influence des teneurs en nutriments sur la croissance des lentilles d’eau (Lemna minor)


Visualisation(s): 3314 (38 ULiège)
Téléchargement(s): 257 (4 ULiège)

Apport du traitement d’images dans le suivi de l’influence des teneurs en nutriments sur la croissance des lentilles d’eau (Lemna minor)

Notes de la rédaction
Reçu le 18 janvier 2013, accepté le 8 octobre 2013
Résumé
Le tapis végétal formé par les lentilles d’eau (Lemna minor) sur le plan d’eau constitue un compartiment important dans les mécanismes épuratoires des bassins de traitement des eaux usées par macrophytes flottants. Une biomasse excessive ou déficitaire serait à la base de certains dysfonctionnements dans ce type de bassins. La modélisation de ces bassins à lentilles d’eau par l’approche de bilan de matière basée sur une « matrice de Petersen » faciliterait une gestion optimale de ce type d’installations. La présente étude vise à évaluer l’efficacité et les limites du traitement d’images dans le suivi de l’influence sur la croissance de Lemna minor des teneurs en azote et phosphore. Les expériences ont été réalisées dans une chambre de cultures (phytotron) à l’aide d’un pilote constitué de cuves cubiques en plexiglas dans lesquelles ont été placées une biomasse fraiche (1 g) de Lemna minor et différentes concentrations d'azote (de 5 à 64 mg N-NH4+·l-1) et de phosphore (de 1 à 24 mg P-PO43-·l-1). L’analyse d’images numériques a été utilisée en complément aux méthodes gravimétriques (masses humide et/ou sèche) pour suivre et quantifier la biomasse. Les résultats ont montré que le traitement d’images numériques constitue une méthode non destructive permettant un suivi et une estimation continus de la biomasse au cours de l’expérience (bonnes corrélations entre surface couverte et masse humide/sèche). La croissance de Lemna minor chute progressivement lorsque les teneurs en azote et en phosphore sont respectivement supérieures à 16 mg N-NH4+·l-1 et 6 mg P-PO43-·l-1. Les taux de croissance maximum (µmax) sont de l’ordre de 0,07 par jour, les constantes cinétiques de demi-saturation (KS) et d’inhibition (KI) valent respectivement 3,83 mg·l-1 et 204 mg·l-1 pour l’azote ainsi que 1,26 mg·l-1 et 13,3 mg·l-1 pour le phosphore. Ces résultats sont intéressants et pourraient servir à la gestion optimale du lagunage par lentilles d’eau et à la modélisation des réacteurs biologiques.
Abstract
Contribution of image processing in monitoring the influence of nutrients on the growth of duckweed (Lemna minor). The growth of duckweed (Lemna minor) is an important component in the treatment process in wastewater floating macrophyte ponds. An excess or shortage of this biomass may be responsible for the dysfunction of such ponds. Modeling these duckweed ponds through mass balances based on Petersen’s matrix may be helpful in facilitating the optimal management of such facilities. This study sought to assess the efficiency of digital image processing in the growth monitoring of Lemna minor under different concentrations of nitrogen and phosphorus. The experiments were carried out in a growth chamber (phytotron) using an experimental pilot involving initial fresh Lemna minor biomass (1 g), nitrogen and phosphorus concentrations varying between 5 and 64 mg N-NH4+·l-1, and 1 and 24 mg P-PO43-·-1, respectively. Digital image processing was achieved in addition to gravimetric methods (fresh weight and/or dry weight). Our results showed that the image processing method allowed a continuous and non-destructive monitoring of duckweed biomass. The growth of Lemna minor progressively decreased when nitrogen and phosphorus concentrations were greater than 16 mg N-NH4+·l-1 and 6 mg P-PO43-·-1. The maximum growth rates μmax were of the order of 0.07 d-1. The kinetic constants KS and KI were respectively 3.83 mg·l-1 and 204 mg·l-1 for nitrogen, and 1.26 mg·l-1 and 13.3 mg·l-1 for phosphorus. Such results are interesting as they could contribute to the optimal management of Lemna minor and the modeling of biological reactors.
1Les diverses activités anthropiques liées aux fortes densités de populations dans les milieux ruraux et urbains entrainent une augmentation de la consommation d'eau et une production importante d'eaux usées. Ces eaux usées nécessitent une bonne gestion afin de réduire d'une part les risques sanitaires pour la population et d'autre part, préserver les écosystèmes récepteurs. Par ailleurs, les conditions climatiques, notamment la température et l'ensoleillement dans ces milieux, permettent d'envisager différentes technologies d'épurations des eaux usées dont le lagunage à macrophytes (Hilman, 1961 ; Oron et al., 1986 ; Zirschky et al., 1988 ; Boniardi et al., 1994 ; Cross, 2002 ; Dalu et al., 2003 ; Monette et al., 2006 ; Lasfar et al., 2007).
2Les lentilles d'eau, Lemna minor, l’espèce la plus courante (Hilman, 1961 ; Cross, 2002), font partie des plantes aquatiques généralement utilisées pour le traitement des eaux résiduelles domestiques ou industrielles du fait de leur performance dans l'élimination de la pollution carbonée et leurs capacités d'assimilation de l'azote ou du phosphore (Debusk et al., 1987 ; Vermaat et al., 1998 ; Al-Nozaily et al., 2000a ; Al-Nozaily et al., 2000b; Koné, 2002). Cependant, leur croissance rapide dans les conditions optimales (i.e. température, intensité lumineuse, ensoleillement, nutriments, pH, etc.) exige une gestion du tapis végétal formé sur le plan d'eau en vue d'assurer une bonne épuration des eaux usées (Filbin et al., 1985 ; Radoux et al., 1992 ; Bonomo et al., 1997 ; Korner et al., 1998; Monette et al., 2006 ; Demirezen et al., 2007 ; Lasfar et al., 2007). Dans cette optique, les lentilles d’eau doivent être régulièrement récoltées. Dans le cas contraire, le tapis végétal devient trop important, les lentilles se chevauchent et leur mortalité augmente. Les lentilles mortes décantent éventuellement et se décomposent au fond du bassin, créant ainsi une nouvelle source de pollution (Debusk et al., 1981 ; Reddy et al., 1983 ; Körneer et al., 1998 ; Jupsin et al., 2004). En outre, une récolte hasardeuse des lentilles pourrait entrainer la diminution de la capacité d’épuration et favoriser le développement d’algues, d’où l’intérêt de suivre et de prédire leur croissance en fonction des paramètres du milieu (luminosité, température, pH, azote, phosphore, alcalinité, DCO, etc.).
3De nombreuses études de suivi et de mesure de la biomasse des lentilles d'eau ont été réalisées principalement sur base de la masse sèche ou humide (Edwards et al., 1992 ; Köner et al., 1998 ; Vermaat et al., 1998 ; Rahmani et al., 1999 ; Caicedo et al., 2000 ; Cedergreen et al., 2002). Toutefois, ces deux méthodes présentent des inconvénients qui ne permettent pas de suivre de manière continue la biomasse. Si la première (masse sèche) est plus fiable mais destructive, la deuxième (masse humide) est par contre non destructive mais moins précise et reproductible. Il existe, par ailleurs, une troisième méthode de mesure de la biomasse des lentilles d'eau basée sur la détermination du nombre de frondes des lentilles d’eau (Lemon et al., 2000). Cette méthode est encore beaucoup moins précise que les précédentes, car elle renseigne davantage sur la reproduction des lentilles d’eau plutôt que sur leur masse, leur croissance ou encore leur mortalité.
4Pour pallier aux inconvénients des méthodes conventionnelles, la présente étude s’intéresse à l’évaluation de l’efficacité et des limites du traitement d’images numériques comme méthode complémentaire aux méthodes gravimétriques (masses sèche et/ou humide) dans le suivi de la biomasse de Lemna minor. Cette méthode présente l’avantage d’être non destructive, quand bien même elle exige une bonne dextérité dans la prise des images et dans l'utilisation du logiciel du traitement d’images (Jupsin et al., 2004). La corrélation surface–biomasse a été établie à cet effet en vue d’apprécier la relation masse (sèche/humide) et surface couverte.
5Par ailleurs, la modélisation par l’approche de bilan de matière basé sur une « matrice de Petersen » des bassins du lagunage par lentilles d'eau pourrait constituer un outil de gestion optimale de ce type d’installations, comme c’est le cas des autres systèmes tels que les boues activées (Henze et al., 1987 ; Jupsin et al., 2003). Pour parvenir à un modèle mathématique du lagunage par lentilles d’eau qui tient compte de l’élimination de l’azote, du phosphore, ainsi que la libération de l’oxygène et la consommation du gaz carbonique par ces plantes, la connaissance des paramètres cinétiques et stœchiométriques qui décrivent le système s’avère être nécessaire. Ainsi, l’impact des teneurs en nutriments (azote et phosphore) sur la croissance de Lemna minor sous une luminosité et température constantes a été mis en évidence dans cette étude à travers les paramètres cinétiques µmax, KS et KI. Les résultats obtenus pourraient servir à la gestion optimale du lagunage par lentilles d’eau et éventuellement à la modélisation des réacteurs biologiques.
6Les lentilles d’eau Lemna minor utilisées dans cette étude ont été récoltées dans un bassin de lagunage de la station d’épuration de Bertrix dans la province du Luxembourg en Belgique. Elles ont été conservées dans des bacs en plastique contenant de l’eau du milieu de la récolte (eaux usées d’origine).
7Les essais de croissance ont été réalisés dans un phytotron (chambre de cultures) en utilisant un pilote constitué de six cuves cubiques ouvertes en plexiglas transparent (12 cm² x 12 cm²). Six séries ont été suivies à raison d’une série1 par semaine (5 jours ouvrables) et définies de manière à suivre l’influence de faibles et fortes teneurs en nutriments sur la croissance de Lemna minor. La luminosité dans le phytotron, mesurée à l’aide d’un luxmètre de marque Skye (310 µmole·m-²·s-1), a été assurée par des lampes à sodium haute pression (400 watts) avec une photopériode de 16 h jour et 8 h nuit. La constance de l’intensité lumineuse a induit une température moyenne du milieu d’environ 28 °C dans chacune des séries. Avant la mise en culture, la biomasse constituée de Lemna minor a été lavée avec de l’eau du robinet à l’aide d’une passoire en vue d’éliminer les impuretés (Vatta et al., 1995 ; Lasfar et al., 2007). Cette biomasse fraiche (1 g) a été successivement mise en culture dans chaque cuve du pilote, dans 500 ml d’eau distillée contenant différentes concentrations en azote et phosphore (Tableau 1). Ces teneurs en nutriments sont obtenues par addition des quantités déterminées de chlorure d’ammonium (NH4Cl) pour l’azote et de monohydrogeno phosphate de sodium dihydraté (Na2HPO4.2H2O) pour le phosphore. Le pH, mesuré in situ, avec un pH-mètre WTW à affichage numérique, était quasi neutre pour l’ensemble des séries. Les pertes d’eau causées par l'évapotranspiration des plantes étaient compensées quotidiennement par ajout d’eau distillée en vue d’empêcher une éventuelle mortalité des plantes (Jupsin et al., 2004).
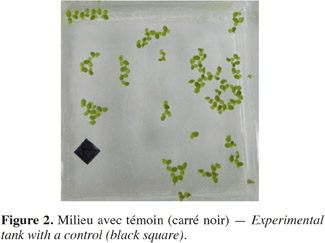
8Le suivi et la mesure de la biomasse des lentilles d'eau mises en culture ont été faits successivement par l’évaluation de la masse humide (MH) et sèche (MS) ainsi que par l’analyse d’images numériques au moyen des logiciels ACD-See® (prétraitement) et Image Pro-Plus® (traitement). Pour la MH, les lentilles prélevées à l’aide d’un tamis de cuisine étaient étalées sur un papier absorbant pendant 5 min et pesées. La MS a nécessité le séchage à l’étuve à 105 °C durant 24 h des lentilles d’eau dont la MF était préalablement mesurée, et puis pesée. La prise des images a été réalisée à l’aide d'un appareil photo numérique compact de marque Nikon® COOLPIX L120, à objectif intégré avec une distance focale de 25-525 mm et ayant une résolution de 14 méga pixels (14 MP) (Tableau 2).


9Le prétraitement a consisté à rogner l’image pour n’obtenir que la surface couverte par les plantes (Figure 2). Le traitement a consisté en un comptage et à la détermination des caractéristiques géométriques de Lemna minor (surface, grand axe, petit axe, périmètre). L’option mode manuel du logiciel par reconnaissance automatique de la couleur de la plante (verte : Lemna vivante) a été utilisé à cet effet. Ce mode permet le comptage automatique de l’objet sur la base de sa couleur sélectionnée par l’utilisateur du logiciel. Ainsi, les lentilles d’eau vertes (vivantes) sont comptées (délimitation en rouge) sur l’image (Figures 3). Les intensités digitales « Red-Green-Blue (RGB) » correspondant aux couleurs des lentilles vivantes étaient comprises entre 100 et 255. Les caractéristiques des lentilles d’eau comptées (i.e. surface, grand axe, petit axe, périmètre) sont générées automatiquement après analyse. Un exemple de ces caractéristiques est donné au tableau 3.
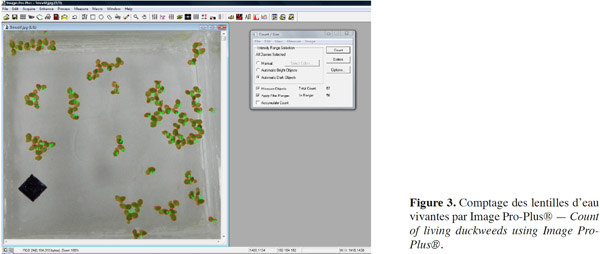

10La corrélation surface-biomasse a été établie en vue d’apprécier la relation masse (sèche/humide) et surface couverte. Pour ce faire, différentes (12) MH connues des lentilles d’eau ont été placées dans différentes cuves ne contenant que 500 ml d'eau distillée. Elles ont été photographiées et ensuite prélevées en conformité avec le protocole afin d’accéder successivement à leurs surfaces couvertes et leurs MS. Les équations (1) et (2) des droites d'étalonnage ont été ainsi écrites :
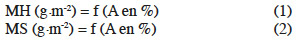
11où A désigne la surface couverte en pourcentage. Cette surface est obtenue en faisant le rapport entre la surface réelle couverte par les lentilles d’eau dans une cuve et la surface totale de la cuve vide (144 cm2).
12Le taux de croissance µ expérimental a été déterminé par régression de la droite lnX = µt + b avec X désignant la biomasse végétale présente et t exprimant l’évolution temporelle de l'expérience. On suppose ainsi une croissance exponentielle de la biomasse qui obéit à la cinétique de Monod (équation 3) (Boniardi et al., 1994 ; Vatta et al., 1995) et qui connait une inhibition par excès de substrat (azote) suivant ainsi une cinétique d’Andrews (équation 4) (Caicedo et al., 2000 ; Jupsin et al., 2004) :
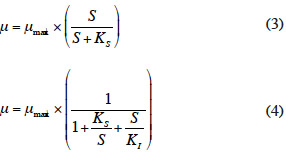
13où µmax, S, KS et KI sont respectivement le taux de croissance maximum (par jour), la teneur en substrats (mg·l-1), la constante de demi-saturation (mg·l-1) et la constante d’inhibition (mg·l-1).
14Par conséquent, les méthodes d’ajustement linéaire (Lineweaver-Burk [L-B], Eadie-Hofstee [E-H], Hanes-Wolff [H-W ou Langmuir]) et non linéaire (algorithme de Gauss-Newton) par moindres carrés ont été respectivement appliquées aux équations (3) et (4) pour déterminer le taux de croissance maximum µmax ainsi que les constantes cinétiques de saturation KS et d’inhibition KI (Fletcher, 1987 ; Henderson, 1992 ; Doran, 1995). Dans les méthodes linéaires, seules les valeurs issues de la méthode qui donnait la plus faible somme des carrés des résidus [∑R² = (µexp-µaj)2] ont été retenues.
15Par la suite, les différentes séries expérimentales ont été globalisées en vue d’obtenir des paramètres cinétiques globaux qui serviraient dans l’équation finale de modélisation de la cinétique de croissance des lentilles d’eau qui, selon Michaelis-Menten2 cité par Lasfar et al. (2007), s’écrit (équation 5) :
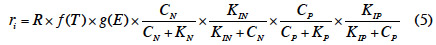
16où ri : taux de croissance (µ) ; R : constante qui s’apparente au taux de croissance maximum (µmax) ; f(T) et g(E) : fonction tenant compte respectivement de l’influence de la température et de la photopériode dans la croissance.
17Les résultats montrent une bonne corrélation entre la surface couverte et la masse (sèche/humide). La valeur de R² tend vers l’unité (Figure 4), ce qui permet de considérer l’analyse d’images comme une méthode fiable d’estimation de la biomasse dans les conditions du test.
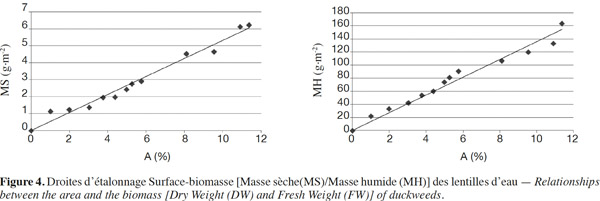
18Il ressort de ces droites d'étalonnage les relations surface-biomasse ci-après :
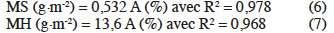
19De ces équations, pour une surface couverte d’environ 10 %, on obtient dans les conditions expérimentales testées, au minimum 136 g MH·m-2 et 5,3 g MS·m-2. Ces équations constituent une information méthodologique intéressante dans l’étude de la cinétique de croissance des lentilles d’eau au laboratoire ou encore dans les conditions de terrain.
20Bien que les présentes valeurs de R2 tendent toutes vers l’unité, les valeurs des pentes sont distinctes par rapport à celles découlant de travaux antérieurs (Jupsin et al., 2004). En effet, ces auteurs ont obtenu respectivement des valeurs de pente de 0,269 et 2,80 pour les relations 6 et 7. Ces écarts entre les valeurs de pentes peuvent s’expliquer par la différence du logiciel principal de traitement d’images, à savoir Image Tools® (Jupsin et al., 2004), qui exige de transformer l’image en niveaux de gris (perte de la qualité de l’image) en vue de compter les objets et déterminer les autres paramètres de taille (i.e. surface). Par contre, cette étape n’existe pas dans le logiciel Image Pro-Plus® où les objets sont comptés et caractérisés juste après la calibration de l’image. En outre, l'espèce expérimentale étant la même, Lemna minor, les milieux où elle se développe sont par contre différents (pauvre ou riche en nutriments), ce qui influencerait l'épaisseur de la plante et la taille des racines (Hilman, 1961 ; Leng, 1999 ; Cedergreen et al., 2002). Il s'ensuit ainsi que la même surface, le même pourcentage de recouvrement des Lemna ne traduirait pas toujours la même quantité de la biomasse (même masse) selon les milieux de récolte et que la calibration devra donc être spécifique au cas étudié. Toutefois, plus la surface couverte augmente, moins la méthode de traitement d’images reste fiable, du fait des possibles recouvrements des lentilles qui sont non statiques dans l’eau (dû à leur croissance). Il serait alors intéressant de travailler dans un milieu où les lentilles ont été diluées pour en diminuer leur densité surfacique. Jupsin et al. (2004) préconise d’ailleurs qu'au-delà d'une certaine densité de Lemna (environ 60 % de couverture), il est nécessaire soit de répartir la biomasse dans deux cuves ou plus afin de photographier chacune d'elles et sommer leurs surfaces, soit de l'introduire dans une plus grande cuve qui subira alors les mêmes prise et traitement de l'image.
21En pratique, la réalisation de l'analyse photographique sur une lagune nécessiterait de prélever un échantillonnage le plus représentatif possible, car il ne sera pas aisé de photographier tout un bassin de lagunage. Cependant, un échantillonnage de quantité (récolte) peut rendre la méthode destructive, d'où l'importance d'un échantillonnage de qualité qui permettrait de minimiser la biomasse prélevée et de refléter le mieux possible la réalité. Par ailleurs, à moins d'améliorer le logiciel ou d'en trouver un plus adéquat (Software duckweed, par exemple), il ne sera pas possible de prendre les photogaphies in situ du fait de l'éclairage (reflet sur l'image) et de la nécessité d'avoir un fond clair et uniforme.
22Séries expérimentales. La figure 5 représente respectivement l'évolution de la surface végétale couverte au cours du temps ainsi que l’évolution du taux de croissance en fonction de la teneur initiale en substrat dans les six séries expérimentées.
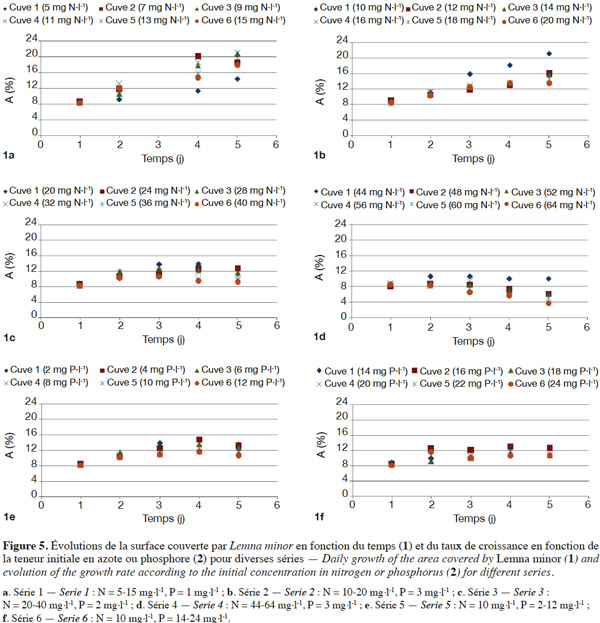
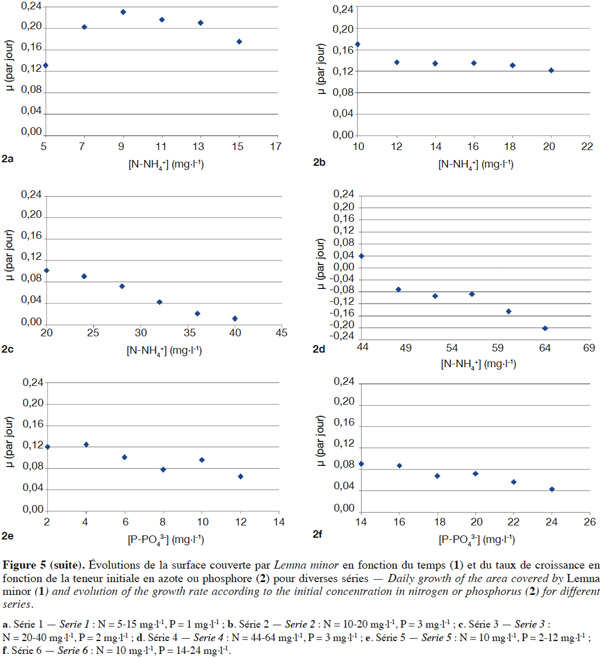
23Ces résultats montrent qu’avec l'azote comme substrat (séries 1, 2, 3, 4), à une luminosité constante de 310 µmole·m-²·s-1, une température de l’ordre de 28 °C et un pH quasi neutre, la biomasse a tendance à croître dans les trois premières séries. Les valeurs de µ sont en effet positives (Figure 5 : 1a, 1b, 1c et 2a, 2b, 2c). Le taux de croissance expérimental le plus élevé est de 0,23 par jour avec des teneurs respectives en azote et en phosphore de 9 mg·l-1 et 1 mg·l-1. Cependant, avec l’augmentation progressive de la teneur en azote (≥ 16 mg N-NH4+·l-1, figure 5 : 2b), il y a une diminution partielle de cette croissance en accord avec Caicedo et al. (2000), Jupsin et al. (2004), Monette et al. (2006) et Lasfar et al. (2007). Cette décroissance tend à devenir sévère à partir de 48 mg N-NH4+·l-1 jusqu’à provoquer la mortalité constatée par le blanchissement de certaines plantes dans les cuves. Des valeurs négatives de µ sont alors obtenues (Figure 5 : 2d). Le taux le plus bas est de - 0,20 par jour, avec des teneurs respectives en azote et en phosphore de 64 mg·l-1 et 2 mg·l-1. Dans le cas du phosphore comme substrat (séries 5, 6) et dans les mêmes conditions expérimentales, des valeurs positives de µ sont obtenues (croissance de la biomasse, figure 5 : 2e et 2f). Néanmoins, ces taux de croissance sont faibles comparativement au cas de l'azote pour des concentrations similaires. La chute de la croissance est observée à de très faibles teneurs (dès 4 mg P-PO43-·l-1, série 5) (Figure 5 : 2e). Le taux de croissance le plus élevé s’établit à 0,12 par jour (10 mg N-NH4+·l-1 et 4 mg P-PO43-·l-1), tandis que le taux le plus bas observé est de 0,044 par jour (10 mg N-NH4+·l-1 et 24 mg P-PO43-·l-1).
24Ces résultats s’expliqueraient par le fait que la croissance de Lemna est optimale dans les milieux pauvres en nutriments, notamment en azote et en phosphore, soit moins de 10 mg N·l-1 et moins de 2 mg P·l-1 (Landolt, 1986). En outre, l’étude de la croissance des lentilles d’eau étant faite en fonction de la teneur initiale en nutriments (azote et phosphore), le non-renouvellement du milieu et l’absence des oligo-éléments dans les milieux de culture expliqueraient la croissance limitée observée des plantes. Elles peuvent alors empêcher le développement de Lemna minor, voire même provoquer la mortalité dans le milieu. Or d’après Oron et al. (1989), cette hypothèse est plus probable quand les temps de séjour sont élevés (> 10 jours). Ainsi, dans notre étude, avec un temps de séjour faible (4 jours), l’hypothèse de la carence en nutriments a été confirmée par les calculs de bilan de matière en substrats (N et P). Cette carence est observée pour des concentrations inférieures ou égales à 28 mg·l-1 et 6 mg·l-1, respectivement pour l’azote et le phosphore. Toutefois, à partir de 32 mg N-NH4+·l-1 et 8 mg P-PO43-·l-1, un excès des nutriments était noté, excès qui pourrait être responsable d’une éventuelle inhibition de la croissance de Lemna minor dans le milieu.
25La cinétique de croissance de Lemna minor a été caractérisée par la détermination des paramètres cinétiques µmax, KS et KIpour chacune des séries. Ces paramètres ont été déterminés par les méthodes de linéarisation et de l'ajustement non linéaire (Tableau 4).
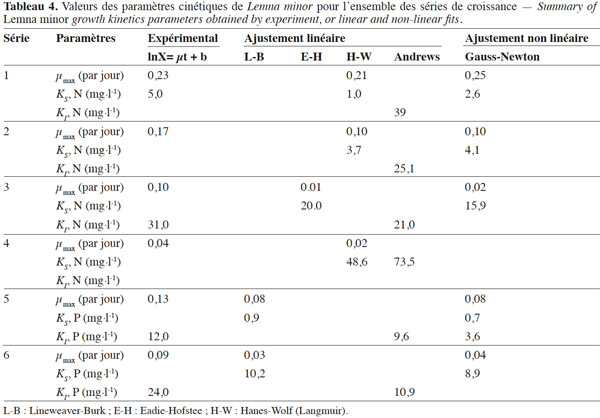
26Malgré des valeurs adéquates de photopériode (16 h/8 h) (Landolt et al., 1987), de température (28 °C) (Oron et al., 1989 ; Boniardi et al., 1999) et d’intensité lumineuse (310 µmole·m-²·s-1) pour une bonne croissance des lentilles d’eau (Filbin et al., 1985 ; Lasfar et al., 2007), les valeurs de µmax ajustées ainsi que celles des constantes cinétiques KS et KI obtenues sont faibles. Ceci est en accord à la fois avec l’hypothèse des concentrations limitantes à des teneurs relativement faibles en nutriments (et en oligo-éléments) et l’hypothèse d’inhibition à des concentrations élevées en nutriments dans le milieu. L’expression globale de la cinétique de croissance s’écrirait ainsi :
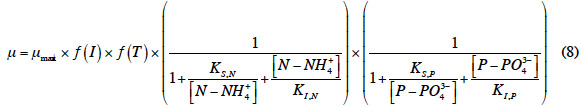
27Globalisation des séries expérimentales. La globalisation de quatre séries expérimentales (1, 2, 3, 4) relatives à l’azote comme substrat en une série confirme les observations selon lesquelles les lentilles d’eau croissent à de faibles teneurs en azote (< 10 mg N·l-1) (Landolt, 1986). La limitation de cette croissance suite à la carence des nutriments et des oligo-éléments est observée jusqu’aux environs de 28 mg N-NH4+·l-1. Cependant, au-délà de cette concentration, l’excès d’azote dans le milieu a tendance à inhiber davantage la croissance de Lemna minor, tel que montré par le bilan des matières (Figure 6a).
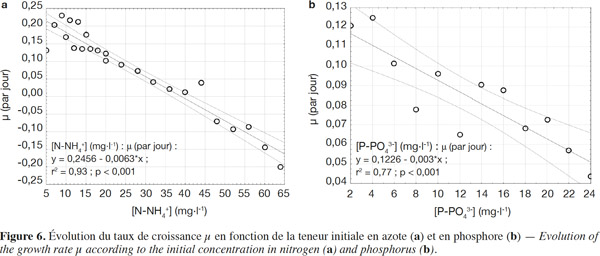
28Pour les séries 5 et 6 relatives au phosphore, les observations sont similaires au cas de l’azote. La tendance à la décroissance est observée dès 6 mg P-PO43-·l-1. D’une manière générale, une augmentation de la teneur en phosphore entrainerait une diminution du taux de croissance µ (Figure 6b). La carence en nutriments est précoce (≤ 6 mg P-PO43-·l-1) et l’excès en phosphore dans le milieu est confirmé à partir de 8 mg P-PO43-·l-1.
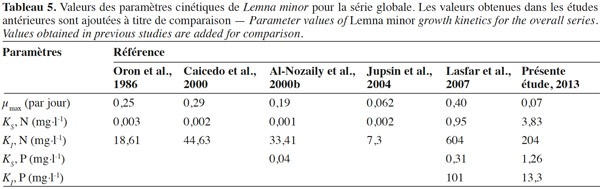
29Il se dégage des paramètres cinétiques obtenus de ces séries globales que, malgré de faibles valeurs de µmax, celles-ci restent néanmoins dans la gamme des valeurs proposées dans la littérature (Tableau 5). Quant aux valeurs des constantes cinétiques, KS, elles sont supérieures à la gamme rencontrée dans la littérature, ce qui n’est pas le cas pour KI. Le milieu n’étant pas renouvelé, il engendre une augmentation de KI et une diminution de KS. Toutefois, ayant très peu de points dans la zone de croissance (< 10 mg N-NH4+·l-1, < 4 mg P-PO43-·l-1, Figure 6), les valeurs de KS issues de cette globalisation ne sont donc pas particulièrement très fiables.
30Dans cette étude, l’efficacité et les limites du traitement d’images comme méthode complémentaire dans le suivi de l’impact des teneurs en nutriments (azote, phosphore) sur la croissance de Lemna minor a été évaluée. Malgré les limites basées sur les corrélations empiriques de la méthode (essai sur pilote), le traitement d’images a l’avantage d’être non destructif dans le suivi continu de la biomasse. Il permet en effet de mesurer la biomasse de par la corrélation surface-masse (humide/sèche). Cependant, un milieu très dense dominé par un recouvrement des plantes exigerait une dilution et une augmentation de surface afin d’aboutir à de bons résultats. L’idéal serait ainsi d’examiner l’aspect de densité surfacique avec cette méthode afin de prévenir les difficultés qu’on rencontrerait en réalité sur terrain (pleine échelle). Les résultats obtenus du point de vue de l’enlèvement des nutriments ne tiennent donc compte que de la phase de la croissance. Ainsi, des expériences menées avec une luminosité de 310 µmole·m-²·s-1 induisant une température d’environ 28 °C, un pH presque neutre, il en résulte que la biomasse de Lemna minor croît faiblement à des teneurs faibles et moyennes en nutriments (azote et phosphore) dans un milieu non renouvelé et exempt d’oligo-éléments. Cette croissance connait une chute progressive à partir de 16 mg N-NH4+·l-1 et de 6 mg P-PO43-·l-1 suite à la limitation des teneurs en nutriments et une possible inhibition due à un excès de substrats à des teneurs élevées. Toutefois, il serait intéressant de faire varier l’ensemble des paramètres (température, intensité lumineuse, photopériode, pH, etc.) qui sont liés à la croissance des lentilles d’eau en vue de mieux cerner l’influence de chacun d’eux dans cette croissance et éventuellement évaluer le taux de mortalité.
31Remerciements
32Ce travail a été réalisé au Département des Sciences et Gestion de l’Environnement de l’Université de Liège et s’inscrit dans le cadre du projet de recherche doctorale du premier auteur. Ce projet de recherche doctorale a été financé par la Coopération Technique Belge (CTB). Nos remerciements au Docteur Louis Amani Kouadio du Lethbridge Research Center (Agriculture and AgriFood Canada) ; aux membres de l’Unité Assainissement et Environnement du Campus d’Arlon de l’Université de Liège ; au personnel du laboratoire d’Écotoxicologie, Management de la Qualité de l’Eau et Biotechnologie environnementale de l’Université de Kinshasa et au personnel du laboratoire de Physique des sols et Hydrologie du Commissariat Général à l’Énergie Atomique/Centre d’Études nucléaires de Kinshasa pour leur collaboration.
Bibliographie
Al-Nozaily F., Alaerts G. & Veenstra S., 2000a. Performance of duckweed-covered sewage lagoons–I. Oxygen balance and COD removal. Wat. Res., 34(10), 2727-2733.
Al-Nozaily F., Alaerts G. & Veenstra S., 2000b. Performance of duckweed-covered sewage lagoons–II. Nitrogen and phosphorus balance and plant productivity. Wat. Res., 34(10), 2734-2741.
Boniardi N., Vatta G., Nano G. & Carra S., 1994. Removal of water polluants by Lemna gibba. Chem. Eng. J., 54, 41-48.
Boniardi N., Rota R. & Nano G., 1999. Effect of dissolved metals on the organic load removal efficiency of Lemna gibba. Wat. Res., 33(2), 530-538.
Bonomo L., Pastorelli G. & Zambon N., 1997. Advantages and limitations of duckweed-based wastewater treatment systems. Wat. Sci. Technol., 35(5), 239-246.
Caicedo J.R., Van Der Steen P., Arce O. & Gijzen H., 2000. Effect of total ammonium nitrogen concentration and pH on growth rates of duckweed (Spirodela polyrrhiza). Wat. Res., 34(15), 3829-3835.
Cedergreen N. & Madsen T.V., 2002. Nitrogen uptake by the floating macrophyte Lemna minor. New Phytol., 155, 285-292.
Cross J.W., 2002. The charms of duckweed, http://www.mobot.org/jwcross/duckweed/duckweed.htm, (15/11/2011).
Dalu J.M. & Ndamba J., 2003. Duckweed based wastewater stabilization ponds for wastewater treatment (a low cost technology for small urban areas in Zimbabwe). Phys. Chem. Earth Parts A/B/C, 28(20-27), 1147-1160.
Debusk T.A., Ryther J.H. & Hanisak M.D., 1981. Effects of seasonality and plant density on the productivity of some freshwater macrophytes. Aquat. Bot., 10, 133-142.
Debusk T.A. & Reedy K.R., 1987. BOD removal in floating aquatic macrophytes based wastewater treatment. Wat. Sci. Technol., 19(12), 273-279.
Demirezen D., Aksoy A. & Uruç K., 2007. Effect of population density on growth, biomass and nickel accumulation capacity of Lemna gibba (Lemnaceae). Chemosphere, 66(3), 553-557.
Doran P.M., 1995. Bioprocess engineering principles. London: Academic Press Limited.
Edwards P., Hassan M.S., Chao C.H. & Pacharaprakiti C., 1992. Cultivation of duckweeds in spetage-loades earthen ponds. Bioresour. Technol., 40, 109-117.
Filbin G.J. & Hough A.R., 1985. Photosynthesis, photorespiration and productivity in Lemna minor L. Limnol. Oceanogr., 30(2), 322-334.
Fletcher R., 1987. Practical methods of optimization. New York, NY, USA: John Wiley & Sons.
Henderson P.J.F., 1992. Statistical analysis of enzyme kinetic data. In: Eisenthal R. & Danson M.J., eds. Enzyme assays: a practical approach. Oxford, UK: IRL Press, 1-58
Henze M. et al., 1987. Activated sludge model No. 1. London: IAWQ.
Hillman W.S., 1961. The Lemnaceae, or duckweed: a review of the descriptive and experimental literature. Bot. Rev., 27, 221-287.
Jupsin H., Praet E. & Vasel J.-L., 2003. Dynamic mathematical model of high rate algal ponds. Water Sci. Technol., 48(2), 197-204.
Jupsin H., Richard H. & Vasel J.-L., 2004. Contribution of floating macrophytes (Lemna sp.) in pond modelization. In: Proceedings of the 6th International Conference on Waste Stabilisation Ponds, September 28th - October 1st, 2004, Avignon, France, 387-393. Antony, France : IRSTEA.
Koné D., 2002. Lagunage à microphytes et à macrophytes (Pistia stratiotes) en Afrique de l'Ouest et du Centre : état des lieux, performances épuratoires et critères de dimensionnement. Thèse de doctorat : École Polytechnique Fédérale de Lausanne, Environnement naturel - Architectural and Construction Faculty (Suisse).
Körner S. & Vermaat J.E., 1998. The relative importance of Lemna gibba, bacteria and algae for the nitrogen and phosphorus removal in duckweed-covered domestic wastewater. Water Res., 32(12), 3651-3661.
Landolt E., 1986. Biosystematic investigations in the family of duckweeds (Lemnaceae). Vol. 2. The Family of Lemnaceae – A monographic study. Vol. 1. Zürich, Switzerland: Veroffentlichungen des Geobotanischen Institutes der Eidgenossischen Technischen Hochschule, Stiftung Rubel.
Landolt E. & Kandeler R., 1987. Biosystematics investigations in the family of duckweeds (Lemnaceae). Vol. 2. The family of Lemnaceae – a monographic study. Vol. 2. Phytochemistry, Physiology, Application, and bibliography. Zürich, Switzerland: Veroffentlichungen des Geobotanischen Institutes der Eidgenossischen Technischen Hochschule, Stiftung Rubel.
Lasfar S., Monette F., Millette L. & Azzouz A., 2007. Intrinsic growth rate: a new approach to evaluate the effects of temperature, photoperiod and phosphorus-nitrogen concentrations on duckweed growth under controlled eutrophication. Water Res., 41, 2333-2340.
Lemon G.D. & Posluszny U., 2000. Comparative shoot development and evolution in the Lemnaceae. Int. J. Plant Sci., 161(5), 733-748.
Leng R.A., 1999. Duckweed: a tiny aquatic plant with enormous potential for agriculture and environment, http://www.fao.org/ag/aga/AGAP/FRG/DW/Dw2.htm, (13/02/2012).
Monette F., Lasfar S., Millette L. & Azzouz A., 2006. Comprehensive modeling of mat density effect on duckweed (Lemna minor) growth controlled eutrophication. Water Res., 40(15), 2901-2910.
Oron G., Porath D. & Wildschut L.R., 1986. Wastewater treatment and renovation by different duckweed species. J. Environ. Eng., 112(2), 247-263.
Oron G. & Willers R.H., 1989. Effects of wastes quality on treatment efficiency with duckweed. Water Sci. Technol., 21, 639-645.
Radoux M. & Kemp D., 1992. Rôle de la fréquence des prélèvements de la biomasse produite sur les capacités épuratrices de Lemna minor L. Rev. Sci. Eau, 5, 55-68.
Rahmani G.N.H. & Sternberg S.P.K., 1999. Bioremoval of lead from water using Lemna minor. Bioresour. Technol., 70, 225-230.
Reddy K.R. & Tucker J.C., 1983. Productivity and nutrient uptake of water hyacinth, Eichhornia crassipes. 1 – Effect on nitrogen source. Econ. Bot., 37(2), 237-247.
Vatta G., Rota R., Boniardi N. & Nano G., 1995. Dynamic modelling of waste-water treatment plants based on Lemna gibba. Chem. Eng. J., 57, 37-48.
Vermaat J.E. & Hanif M.K., 1998. Performance of common duckweed species (Lemnaceae) and waterfern Azolla filiculoides on different types of waste water. Wat. Res., 32(9), 2569-2576.
Zirschky J. & Reed S.C., 1988. The use of duckweed for wastewater treatment. J. Water Pollut. Control Fed., 60(7), 1253-1258.
Notes
1 Série : ensemble de six milieux de culture (azote, phosphore et eau distillée) contenus dans six cuves cubiques constituant le pilote.
2 Michaelis L. & Menten M.L., 1913. Die Kinetik der Invertinwirkung. Biochem Z., 49, 333-369.
