
Habitat diversity of the Multicolored Asian ladybeetle Harmonia axyridis Pallas (Coleoptera: Coccinellidae) in agricultural and arboreal ecosystems: a review
Univ. Liege - Gembloux Agro-Bio Tech. Unit of functional and evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium). E-mail: a.vandereycken@ulg.ac.be
Univ. Liege - Gembloux Agro-Bio Tech. Unit of functional and evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Univ. Liege - Gembloux Agro-Bio Tech. Unit of functional and evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Univ. Liege - Gembloux Agro-Bio Tech. Unit of functional and evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Univ. Liege - Gembloux Agro-Bio Tech. Unit of functional and evolutionary Entomology. Passage des Déportés, 2. B-5030 Gembloux (Belgium).
Received on April 17, 2012; accepted on October 2, 2012
Résumé
Diversité des habitats de la coccinelle asiatique Harmonia axyridis Pallas (Coleoptera : Coccinellidae) au sein des différents écosystèmes agricoles et forestiers (synthèse bibliographique). La coccinelle asiatique, Harmonia axyridis (Pallas), originaire d'Asie, est une espèce invasive dans de nombreux pays européens et américains. Introduite comme agent de contrôle biologique afin de lutter contre les pucerons et les cochenilles dans les serres, cette espèce a rapidement envahi différents habitats tels que les forêts, les cultures agricoles, les prairies, les jardins et les vergers. Cet article présente une synthèse des habitats où H. axyridis a été observée lors d'inventaires et dans lesquels elle a été utilisée dans la gestion de lutte contre les ravageurs. Cent-six taxons de plantes sur lesquelles H. axyridis a été observée (35 espèces arborescentes, 21 espèces de culture, 27 espèces herbacées, 11 espèces ornementales et 12 espèces de verger) et 89 relations plantes-proies (34 espèces arborescentes, 16 espèces de culture, 13 espèces herbacées, 10 espèces ornementales et 16 espèces de verger) ont été dénombrés dans différents pays. Harmonia axyridis est plus abondante en milieu forestier et plus précisément sur Acer, Salix, Tilia et Quercus qu'en milieu agricole. Certaines espèces végétales, telles que l'ortie Urtica dioica L., présentes à proximité des habitats agricoles, contiennent de grandes quantités d'H. axyridis et peuvent donc servir de réserve de prédateurs avant les invasions de pucerons en grandes cultures. Cette synthèse bibliographique souligne la polyphagie et le comportement eurytopique de cette coccinelle exotique.
Abstract
The Multicolored Asian ladybeetle, Harmonia axyridis (Pallas), native to Asia, is an invasive species in many European and American countries. Initially introduced as a biological control agent against aphids and coccids in greenhouses, this alien species rapidly invaded many habitats such as forests, meadows, wetlands, and agricultural crops. This paper reviews the habitats (forests, crops, herbs, gardens and orchards) where H. axyridis has been observed, either during insect samplings or as part of Integrated Pest Management (IPM) programs. Studies have referenced H. axyridis on 106 plant taxa (35 arboreal species, 21 crop species, 27 herbaceous species, 11 ornamental species, and 12 orchard species) and have identified 89 plant-prey relationships (34 arboreal species, 16 crop species, 13 herbaceous species, 10 ornamental species, and 16 orchard species) in different countries. Harmonia axyridis is more abundant in forest areas, principally on Acer, Salix, Tilia and Quercus, than in agroecosystems. Some plant species, such as Urtica dioica L., which surround crops, contain large numbers of H. axyridis and could constitute important reserves of this alien species in advance of aphid invasions into crops. This review highlights the polyphagy and eurytopic aspect of H. axyridis.
1. Introduction
1The Multicolored Asian ladybeetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), is native to south-east Asia, between Siberia and China (Chapin, 1965). This species has long been used as a biological control agent against aphids and coccids on both sides of the Atlantic Ocean. Since 1988, H. axyridis has become established in at least 38 countries around the world: 9 in America, 26 in Europe and 3 in Africa (Brown et al., 2011).
2In America, the first introduction of H. axyridis was conducted in California in 1916, but the first established populations were referenced 72 years later (Gordon, 1985; Chapin et al., 1991). In this area, H. axyridis has been commonly used for biological control in diverse crops such as pecans (Tedders et al., 1994), red pines (McClure, 1987), apple orchards (Brown et al., 1998), soybeans (Fox et al., 2004), sweet corn (Musser et al., 2003), alfalfa (Buntin et al., 1997; Colunga-Garcia et al., 1998), cotton (Wells et al., 2001), tobacco (Wells et al., 1999) and winter wheat (Colunga-Garcia et al., 1998). Harmonia axyridis was introduced into Europe in 1964 and has been commercialized as a biological control agent since 1982 (Iperti, 1991; Katsoyannos et al., 1997). Thanks to a rapid reproductive cycle allowing this beetle to achieve two or three generations per year, H. axyridis has become the dominant coccinellid in many ecosystems (Brown et al., 2008).
3Harmonia axyridis has been released as a biocontrol agent into numerous crops, arboreal habitats, and orchards (Tables 1 to 5). These releases partly explain the rapid expansion of this alien ladybeetle, favored by its notable dispersal abilities (Koch et al., 2006).
4This review will focus on the natural and semi-natural habitats of the multicolored Asian ladybeetle H. axyridis both inside and outside of its native range, and will highlight its associated plant species and prey species.
2. Habitats of Harmonia axyridis
2.1. Arboreal habitats
5In both native and invaded areas, arboreal habitats are commonly colonized by H. axyridis for growth and reproduction. The most common trees on which H. axyridis is observed are Acer, Salix, Tilia, Quercus and Pinus (LaMana et al., 1996; Osawa, 2000; Adriaens et al., 2008; Brown et al., 2008) (Table 1). In Oregon, this species represents 70% of the coccinellid community in forest stands, where it is often considered as a dominant generalist aphidophagous predator (LaMana et al., 1996). In western Europe, Adriaens et al. (2008) inventoried several habitats and found that H. axyridis was present on more than 100 plant species. In that study, 52% of H. axyridis observations were made on trees and 14% on shrubs. The plants on which H. axyridis was most frequently observed were Acer, Salix, Tilia, Quercus, Betula, Pinus, and Crataegus. In eastern Europe, the first detection of H. axyridis was recorded on the same arboreal species, including Tilia, Quercus and Acer, respectively infested by the following aphid species: Eucallipterus tiliae L. (Hemiptera: Aphididae), Myzocallis walshii Monell (Hemiptera: Aphididae) and Drepanosiphum sp. (Hemiptera: Aphididae) (Tomov et al., 2009) (Table 1).

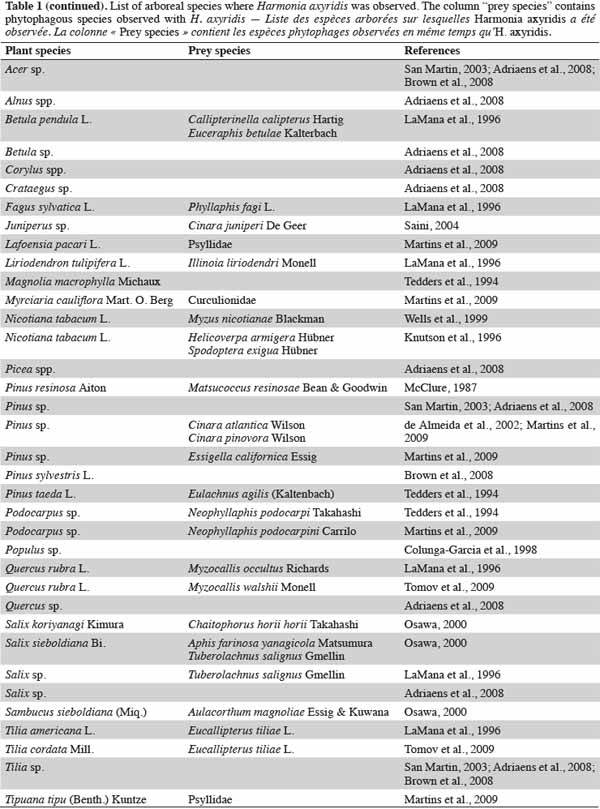
6The presence of forest ecosystems increases the occurrence of H. axyridis in the surrounding habitats such as agroecosystems (Gardiner et al., 2009). A consequence of a higher density of the species in landscapes in close proximity to forests is that predation on native species is higher than that observed in fields surrounded by other croplands (Gardiner et al., 2011).
7Trees are not the only arboreal areas where H. axyridis are able to find food. In addition, H. axyridis has been observed on shrubs, feeding on Tinocallis kahawaluokalani Kirkaldy (Hemiptera: Aphididae) and has also been observed on crape myrtle, Lagerstroemia indica L. (de Almeida et al., 2002).
8Most of these studies have reported observations of H. axyridis mainly in arboreal habitats and in high density. Two reasons may explain the occurrence of this alien coccinellid in arboreal habitats: first, the arboreal taxa cited are affected by high aphid populations in spring and are thus attractive to the coccinellid; second, these arboreal taxa are the most popular taxa in Europe. Further information on studies concerning H. axyridis in arboreal ecosystems can be seen in table 1. Thirty-five plant species and 34 plant-prey-predator relationships have been observed on diverse continents.
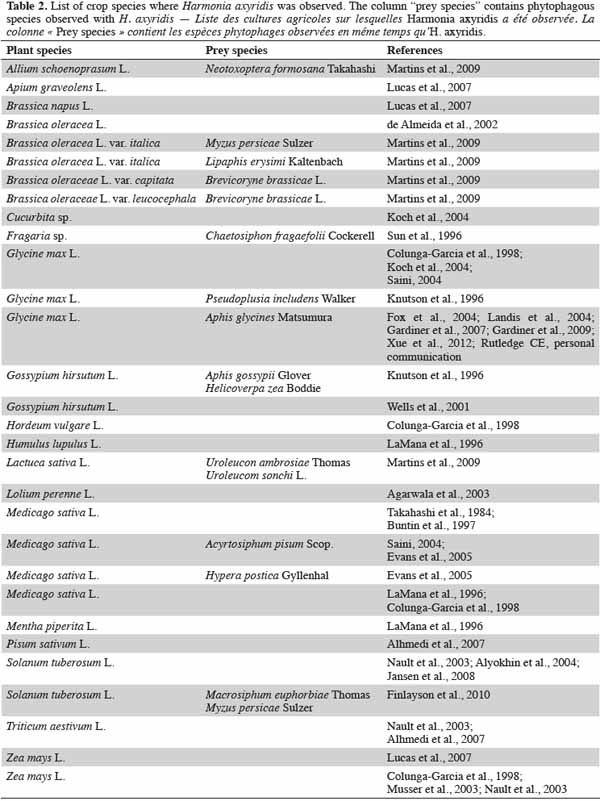
2.2. Agroecosystems
9Agroecosystems may be infested by large quantities of prey and so can constitute habitats where ladybeetles are able to thrive and reach their adult stage. Nevertheless, in these particular ecosystems, the numbers of the most abundant coccinellid species are generally low (three or four dominant species) (Hodek et al., 1996). The composition of the aphidophagous community differs from one crop to another and is influenced by several abiotic factors, such as insolation and humidity (Honěk, 1985) and by biotic factors including the quantity and quality of host plants (Alhmedi et al., 2009), aphid density and diversity (Wright et al., 1980; Honěk, 1982; Thalji, 2006) and adjacent habitats (Colignon et al., 2000; Alhmedi et al., 2009).
10Many studies highlight the dominance of H. axyridis in crop areas, e.g.: in Minnesota, 10 years after its initial detection in 1994 (Koch et al., 2003), H. axyridis rapidly became the most abundant generalist predator in corn, reaching 77.4% of Coccinellidae on the crop (Koch et al., 2006). In other cases, H. axyridis can quickly become the dominant species, as was observed in Michigan crops (alfalfa, soybean, corn and winter wheat), where four years after its arrival, this ladybeetle became dominant, with its proportions varying from 2.8% to 32.3% between 1994 and 1998 on all inventoried crops (Colunga-Garcia et al., 1998). However, some reports claim contrary conclusions. Nault et al. (2003) showed that three years after the arrival of H. axyridis in North Carolina agroecosystems, the most abundant species were not H. axyridis but Coccinella septempunctata L., Coleomegilla maculata De Geer and Hippodamia convergens Guérin-Méneville. In this area, H. axyridis colonized wheat, corn, and potato but reproduced only in wheat and potato (Kidd et al., 1995; Nault et al., 2003). In tobacco plants in Georgia, during 1997 and 1998 (15 years after initial release), H. axyridis was dominated by H. convergens and C. septempunctata (Wells et al., 1999). Finally, in alfalfa in Japan, adult and larval proportions of H. axyridis were found to respectively form 24.6% and 15.5% of coccinellids, figures lower than those of C. septempunctata and Propylea japonica (Thunberg) (Takahashi et al., 1984).
11Variations in coccinellid dominance from year to year or between crops are dependent on multiple factors, but these variations are complex and misunderstood. For example, in agricultural habitats, population densities can be influenced by margin strips of herb species situated in close proximity. Nettle, Urtica dioica L., is known to be a plant species hosting many H. axyridis (Adriaens et al., 2008); the presence of nettle among margin field crops (wheat, green pea) therefore significantly increases the density of H. axyridis (Alhmedi et al., 2007) in these crops. These surrounding areas (nettle, forest) may provide a population of H. axyridis before aphid invasion in the adjacent field crops. The prey species is also an important factor determining the abundance and reproduction rate of H. axyridis. Evans et al. (2005) highlighted that in alfalfa, H. axyridis is an efficient predator, consuming both pea aphids Acyrthosiphon pisum Scop. (Hemiptera: Aphididae) and larvae of alfalfa weevils Hypera postica (Gyllenhal) (Coleoptera: Curculionidae), but that the consumption rates of aphids were greater than those for the weevil. Moreover, when alfalfa weevil is the available food source, H. axyridis does not reproduce, and both larval survival and development of H. axyridis are low (Evans et al., 2005). A second example highlights the problem of prey quality: Uroleucon nigrotuberculatum (Olive) (Hemiptera: Aphididae) on Solidago canadensis L. represent unsuitable prey for H. axyridis during its life cycle (Kamo et al., 2011). These prey species are considered by Hodek et al. (1996) to be “acceptable but inadequate prey” for H. axyridis.
12At the beginning of the 1990s, LaMana et al. (1996) highlighted that H. axyridis was more abundant in arboreal habitats than in agricultural areas. Their results showed that only 4% of H. axyridis were observed in agroecosystems such as alfalfa Medicago sativa L., clover Trifolium sp. L. and peppermint Mentha piperita L. In these ecosystems, H. axyridis was observed in association with 17 aphid species on 17 host plants. While H. axyridis has been more observed in arboreal habitats, it can still thrive and dominate the aphidophagous guild in agroecosystems. In agricultural systems of eastern Canada (pome fruit, grape, field corn, sweet corn, sweet pepper, lettuce, and soybean), H. axyridis was clearly one of the dominant coccinellid species from 1999 to 2003 (Lucas et al., 2007). Unlike in American and Asian studies, H. axyridis presented in Belgium high population levels only one year after its original detection in potato fields (2004-2005) and was able to complete its larval development even with low aphid population, or with no aphids at all (Jansen et al., 2008). In this invaded area, H. axyridis was found to be one of the most abundant coccinellids along with C. septempunctata and Propylea quatuordecimpunctata L. (Jansen et al., 2008).
13One of the most important pests of American soybean is the soybean aphid, Aphis glycines Matsumura (Hemiptera: Aphididae). In 2000, H. axyridis was found to constitute more than 25% of the aphidophagous species in this crop; other such species present included Orius insidiosus (Says) (Hemiptera: Anthocoridae) and Leucopis spp. (Diptera: Chamaemyiidae), which are able to control A. glycines (Fox et al., 2004). Harmonia axyridis was found to cause a reduction in aphid populations by 21-56% in the early season and by 54-95% in midseason (Landis et al., 2004). Five years later, H. axyridis was found to occur at levels of 45 to 62% of the total coccinellid community (Gardiner et al., 2009).
14Despite the fact that H. axyridis is an arboreal species, a high diversity of crops (21) is subject to invasion by this coccinellid (Table 2). Harmonia axyridis invades selected crops according to the aphid species present and to the microclimate inside the field. The high density and high voracity of H. axyridis make it a highly efficient beneficial species. It would be interesting to use H. axyridis as part of a pest control approach (e.g. through a push-pull strategy) in organic crops to control aphid populations.
2.3. Herbs, domestic and ornamental gardens
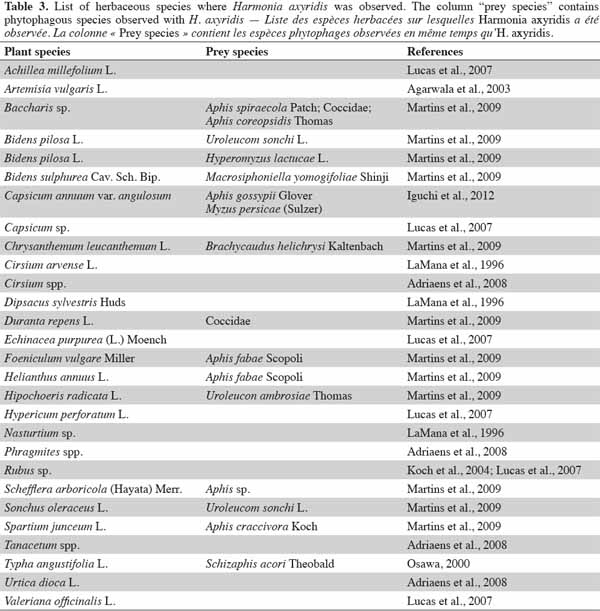
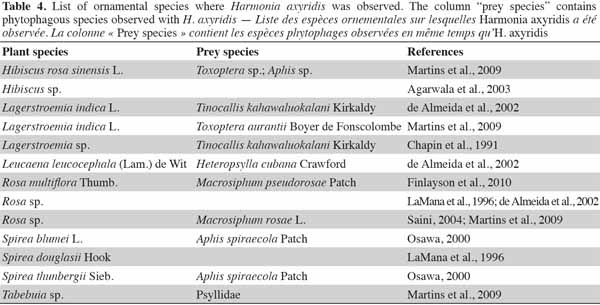
15Harmonia axyridis also occurs in herbaceous habitats such as heathland, meadows, and wetlands (Adriaens et al., 2008). Among these habitats, H. axyridis has been most frequently observed on U. dioica (Adriaens et al., 2008), but also on Bidens pilosa L. (Martins et al., 2009), Phragmites spp. (Adriaens et al., 2008), Artemisia vulgaris L. (Agarwala et al., 2003), and Typha angustifolia L. (Osawa, 2000). The alien species has also been observed in urban habitats, such as in domestic gardens, on Hibiscus rosa sinensis L. (Martins et al., 2009), Lagerstroemia indica L. (Chapin et al., 1991), and Rosa sp. (de Almeida et al., 2002).
16Harmonia axyridis can be used as a biocontrol agent in gardens, but it is not common e.g.: H. axyridis can be used to control Chaetosiphon fragaefolii (Cockerell) (Hemiptera: Aphididae) on strawberry (Sun et al., 1996) and Macrosiphum rosae L. (Hemiptera: Aphididae) on roses (Finlayson et al., 2010). Harmonia axyridis can also be used to control pests on ornamental plant species e.g.: on greenhouse roses (Rosa hybrida L.), the aphid, Macrosiphum euphorbiae Thomas (Hemiptera: Aphididae), can be controlled by H. axyridis and a wasp, Aphelinus asychis Walker (Hymenoptera: Aphelinidae). Harmonia axyridis can be used to complement aphid biocontrol, without disrupting control through intraguild predation (Snyder et al., 2004).
17The presence of H. axyridis in gardens is more anecdotal. The use of ladybeetles as beneficial species needs to be focused on native species such as A. bipunctata. Further examples of H. axyridis habitats, including herbs and ornamental species, are presented in tables 3 and 4. Harmonia axyridis individuals have been observed on herbs and ornamental habitats containing respectively 27 and 11 plant taxa.
2.4. Orchards
18Before winter, coccinellids also thrive in orchards, for example, on apples (Malus spp.) and citrus (Citrus spp.). In these habitats, they accumulate fat content and glycogen reserves (polyol myo-inositol) in response to a decline in food resources (Hodek, 1986; Watanabe, 2002; Hodek, 2011). A consequence of insects feeding on apples and pears is that it causes blemishing of the fruits and reduces the value of the crop (Majerus et al., 2006). In Virginian orchards, H. axyridis has become the most abundant coccinellid species (representing between 40.9% and 90.7% of those species) and it sometimes replaces the dominant native species (Brown, 2003). In citrus orchards (Florida), H. axyridis was also found to be the most abundant coccinellid species between 1997 and 2001. During this period, H. axyridis replaced Cycloneda sanguine (L.), which had been the numerically dominant species before 1997 (Michaud, 2002). In citrus, H. axyridis has also been tested successfully for the control of the root weevil Diaprepes abbreviatus (L.) (Coleoptera: Curculionidae), a major pest in citrus orchards (Stuart et al., 2002). Similar results have been obtained with the citrus aphid, Aphis spiraecola Patch (Hemiptera: Aphididae) and with Toxoptera citricida (Kirkaldy) (Hemiptera: Aphididae) in Florida (Michaud, 2000). In apple orchards in West Virginia, H. axyridis has also provided good biological control of A. spiraecola. However, the use of this alien species as a biocontrol agent has displaced C. septempunctata (Brown et al., 1998). Harmonia axyridis has been used effectively many times in pecan orchards, Carya illinoinensis Wangenh., to control the pecan aphid complex comprising Melanocallis caryaefolia (Davis) (Hemiptera: Aphididae), Monellia caryella Fitch (Hemiptera: Aphididae) and Monelliopsis pecanis Bissell (Hemiptera: Aphididae) (Tedders et al., 1994; LaRock et al., 1996). In Georgia, H. axyridis was released in pecan with legume cover between 1978 and 1981. Nine years after its release, the species had spread and was recorded at a distance of 174 km from the release point. In 1994, H. axyridis was found to be the dominant species at the release site, comprising 54% of coccinellids (Tedders et al., 1994). In Argentina, H. axyridis was also found to be the most abundant coccinellid in Carya sp., with its proportion among coccinellids increasing from 51% in 2001 to 74% in 2003 (Saini, 2004).
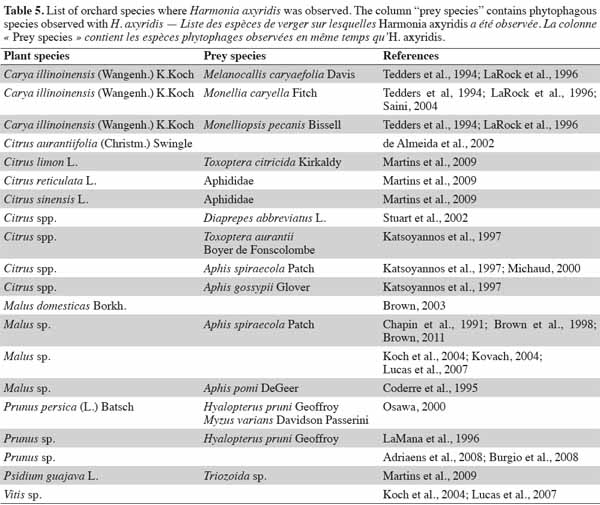
19The presence of H. axyridis in orchard crops is more controversial than in other crops. While H. axyridis acts as efficient biocontrol agent, in the fall season, the species causes damage to fruits. The solution might be to catch H. axyridis adults once the presence of aphids has reached its peak, in order to avoid yield loss through fruit damage. For more examples, see table 5 for a list of studies of H. axyridis in orchard ecosystems. Twelve plant species and 16 plant-prey relationships have been observed on diverse continents.
3. Conclusion
20This review focused on the diversity of natural and semi-natural habitats where H. axyridis has been observed or introduced as a biocontrol agent. The following topics were considered: evaluation of the efficiency of H. axyridis for biological control, the impact of H. axyridis on native species, its occurrence among aphidophagous species, and the evolution of its invasion. Sixty-six studies have referenced H. axyridis on 106 plant taxa (35 arboreal species, 21 crop species, 27 herbaceous species, 11 ornamental species, and 12 orchard species) and have identified 89 plant-prey relationships (34 arboreal species, 16 crop species, 13 herbaceous species, 10 ornamental species, and 16 orchard species) in different countries. This diversity of plants and habitats where H. axyridis has been observed has been used to justify the classification of this alien species as an eurytopic species. The high abundance of H. axyridis within the aphidophagous guild in the majority of the habitats where it is found has had an impact on native species. This highlights the ecological problems provoked by H. axyridis: its presence causes a decline in biodiversity and the displacement of native species. Harmonia axyridis is one of the most abundant species in all natural and semi-natural ecosystems. Its ability to spread across ecosystems raises the interesting proposition of using individuals of the species in nature to control aphid populations. A push-pull strategy could be implemented to control alien species populations in order to decrease their impact on native aphidophagous species.
21Acknowledgements
22This research was funded by the Service Public de Wallonie (SPW – DGO3, project No. D31-1247). Delphine Durieux was financially supported by a PhD grant from the Fonds pour la Formation à la Recherche dans l’Industrie et l’Agriculture (FRIA), Belgium.
Bibliographie
Adriaens T., Gomez G.M.Y. & Maes D., 2008. Invasion history, habitat preferences and phenology of the invasive ladybird Harmonia axyridis in Belgium. BioControl, 53(1), 69-88.
Agarwala B.K., Bardhanroy P., Yasuda H. & Takizawa T., 2003. Effects of conspecific and heterospecific competitors on feeding and oviposition of a predatory ladybird: a laboratory study. Entomol. Exp. Appl., 106(3), 219-226.
Alhmedi A., Haubruge É., Bodson B. & Francis F., 2007. Aphidophagous guilds on nettle (Urtica dioica) strips close to fields of green pea, rape and wheat. Insect Sci., 14, 419-424.
Alhmedi A., Haubruge É. & Francis F., 2009. Effect of stinging nettle habitats on aphidophagous predators and parasitoids in wheat and green pea fields with special attention to the invader Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Entomol. Sci., 12(4), 349-358.
Alyokhin A. & Sewell G., 2004. Changes in a lady beetle community following the establishment of three alien species. Biol. Invasion, 6(4), 463-471.
Brown M.W., 2003. Intraguild responses of aphid predators on apple to the invasion of an exotic species, Harmonia axyridis. BioControl, 48(2), 141-153.
Brown M.W. & Miller S.S., 1998. Coccinellidae (Coleoptera) in apple orchards of eastern West Virginia and the impact of invasion by Harmonia axyridis. Entomol. News, 109(2), 143-151.
Brown P.M.J. et al., 2008. Harmonia axyridis in Great Britain: analysis of the spread and distribution of a non-native coccinellid. BioControl, 53(1), 55-67.
Brown P.M.J. et al., 2011. The global spread of Harmonia axyridis (Coleoptera: Coccinellidae): distribution, dispersal and routes of invasion. BioControl, 56(4), 623-641.
Buntin G.D. & Bouton J.H., 1997. Aphid (Homoptera: Aphididae) management in alfalfa by spring grazing with cattle. J. Entomol. Sci., 32(3), 332-341.
Burgio G., Lanzoni A., Accinelli G. & Maini S., 2008. Estimation of mortality by entomophages on exotic Harmonia axyridis versus native Adalia bipunctata in semi-field conditions in northern Italy. BioControl, 53(1), 277-287.
Chapin E.A., 1965. Coleoptera: Coccinellidae. Insects Micronesia, 16(5), 189-254.
Chapin J.B. & Brou V.A., 1991. Harmonia axyridis (Pallas), the 3rd species of the genus to be found in the United States (Coleopera, Coccinellidae). Proc. Entomol. Soc. Washington, 93(3), 630-635.
Coderre D., Lucas É. & Gagne I., 1995. The occurrence of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae) in Canada. Can. Entomol., 127(4), 609-611.
Colignon P., Hastir P., Gaspar C. & Francis F., 2000. Effets de l’environnement proche sur la biodiversité entomologique en cultures maraichères de plein champ. Parasitica, 56, 59-70.
Colunga-Garcia M. & Gage S.H., 1998. Arrival, establishment, and habitat use of the multicolored Asian lady beetle (Coleoptera: Coccinellidae) in a Michigan landscape. Environ. Entomol., 27(6), 1574-1580.
De Almeida L.M. & da Silva V.B., 2002. First record of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae): a lady beetle native to the Palaearctic region. Rev. Bras. Zoologia, 19(3), 941-944.
Evans E.W. & Gunther D.I., 2005. The link between food and reproduction in aphidophagous predators: a case study with Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol., 102(3), 423-430.
Finlayson C., Alyokhin A., Gross S. & Porter E., 2010. Differential consumption of four aphid species by four lady beetle species. J. Insect Sci., 10.
Fox T.B., Landis D.A., Cardoso F.F. & Difonzo C.D., 2004. Predators suppress Aphis glycines Matsumura population growth in soybean. Environ. Entomol., 33(3), 608-618.
Gardiner M.M. & Landis D.A., 2007. Impact of intraguild predation by adult Harmonia axyridis (Coleoptera: Coccinellidae) on Aphis glycines (Hemiptera: Aphididae) biological control in cage studies. Biol. Control, 40(3), 386-395.
Gardiner M.M. et al., 2009. Landscape composition influences patterns of native and exotic lady beetle abundance. Diversity Distrib., 15(4), 554-564.
Gardiner M.M., O'Neal M.E. & Landis D.A., 2011. Intraguild predation and native lady beetle decline. PLoS ONE, 6(9).
Gordon R.D., 1985. The Coccinellidae (Coleoptera) of America north of Mexico. Journal of the NewYork Entomological Society, 93(1). Lawrence, KS, USA: Allen Press.
Hodek I., 1986. Life cycle strategies, diapause and migration in aphidophagous Coccinellidae. In: Hodek I., ed. Ecology of Aphidophaga. Dordrecht, The Netherlands: Dr W. Junk, 155-166.
Hodek I., 2011. Adult diapause in Coleoptera. Psyche, 2012.
Hodek I. & Honěk A., 1996. Ecology of Coccinellidae. Vol. 54. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Honěk A., 1982. Factors which determine the composition of field communities of adult aphidophagous Coccinellidae (Coleoptera) J. Appl. Entomol., 94(2), 157-168.
Honěk A., 1985. Habitat preferences of aphidophagous coccinellids (Coleoptera). Entomophaga, 30(3), 253-264.
Iguchi M., Fukushima F. & Miura K., 2012. Control of Aphis gossypii and Myzus persicae (Hemiptera: Aphididae) by a flightless strain of Harmonia axyridis (Coleoptera: Coccinellidae) on green pepper plants in open fields. Entomol. Sci., 15(1), 127-132.
Iperti G., 1991. Abiotic and biotic factors influencing distribution of the aphidophagous coccinellidae. In: Polgar L., Chambers R.J., Dixon A.F.G. & Hodek I., eds. Behaviour and impact of Aphidophaga. The Hague, The Netherlands: SPB Academic Publishing, 163-166.
Jansen J. & Hautier L., 2008. Ladybird population dynamics in potato: comparison of native species with an invasive species, Harmonia axyridis. BioControl, 53, 223-233.
Kamo T. & Tokuoka Y., 2011. Influence of the prey aphid Uroleucon nigrotuberculatum parasitizing Solidago canadensis on the larval and adult survivorship of the predatory ladybird beetle Harmonia axyridis. Ecol. Res., 26(2), 471-476.
Katsoyannos P., Kontodimas D.C., Stathas G.J. & Tsartsalis C.T., 1997. Establishment of Harmonia axyridis on citrus and some data on its phenology in Greece. Phytoparasitica, 25(3), 183-191.
Kidd K., Nalepa C., Day E. & Waldvogel M., 1995. Distribution of Harmonia axyridis (Pallas)(Coleoptera: Coccinellidae) in North Carolina and Virginia. Proc. Entomol. Soc. Washington, 97(3), 729-731.
Knutson A.E. & Ruberson J., 1996. Field guide to predators, parasites and pathogens attacking insect and mite pests of cotton: recognizing the good bugs in cotton. College Station, TX, USA: The Texas A&M University System.
Koch R.L. & Hutchinson W.D., 2003. Phenology and blacklight trapping of the multicolored Asian lady beetle (Coleoptera: Coccinellidae) in a Minnesota agricultural landscape. J. Entomol. Sci., 38(3), 477-480.
Koch R.L., Burkness E.C., Burkness S.J.W. & Hutchinson W.D., 2004. Phytophagous preferences of the multicolored Asian lady beetle (Coleoptera: coccinellidae) for autumn-ripening fruit. J. Econ. Entomol., 97(2), 539-544.
Koch R.L., Burkness E.C. & Hutchinson W.D., 2006. Spatial distribution and fixed-precision sampling plans for the ladybird Harmonia axyridis in sweet corn. BioControl, 51(6), 741-751.
Kovach J., 2004. Impact of multicolored Asian lady beetles as a pest of fruit and people. Am. Entomol., 50(3), 159-161.
LaMana M.L. & Miller J.C., 1996. Field observations on Harmonia axyridis Pallas (Coleoptera: Coccinellidae) in Oregon. Biol. Control, 6(2), 232-237.
Landis D.A., Fox T.B. & Costamagna A.C., 2004. Impact of multicolored Asian ladybeetle as a biological control agent. Am. Entomol., 50(3), 153-154.
LaRock D.R. & Ellington J.J., 1996. An integrated pest management approach, emphasizing biological control, for pecan aphids. Southwestern Entomol., 21(2), 153-166.
Lucas É. et al., 2007. The multicolored Asian ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae) in Quebec agroecosystems ten years after its arrival. Eur. J. Entomol., 104(4), 737-743.
Majerus M., Strawson V. & Roy H., 2006. The potential impacts of the arrival of the harlequin ladybird, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), in Britain. Ecol. Entomol., 31(3), 207-215.
Martins C.B.C. et al., 2009. Harmonia axyridis: a threat to Brazilian Coccinellidae? Rev. Brasil. Entomol., 53(4), 663-671.
McClure M.S., 1987. Potential of the Asian predator, Harmonia axyridis Pallas (Coleoptera: Coccinellidae), to control Matsucocus resinosae Bean and Godwin (Homoptera: Margarodidae) in the United-States. Environ. Entomol., 16(1), 224-230.
Michaud J.P., 2000. Development and reproduction of ladybeetles (Coleoptera: Coccinellidae) on the citrus aphids Aphis spiraecola patch and Toxoptera citricida (Kirkaldy) (Homoptera: Aphididae). Biol. Control, 18(3), 287-297.
Michaud J.P., 2002. Invasion of the Florida citrus ecosystem by Harmonia axyridis (Coleoptera: Coccinellidae) and asymmetric competition with a native species, Cycloneda sanguinea. Environ. Entomol., 31(5), 827-835.
Musser F.R. & Shelton A.M., 2003. Bt sweet corn and selective insecticides: impacts on pests and predators. J. Econ. Entomol., 96(1), 71-80.
Nault B.A. & Kennedy G.G., 2003. Establishment of multicolored Asian lady beetle in Eastern North Carolina: seasonal abundance and crop exploitation within an agricultural landscape. BioControl, 48(4), 363-378.
Osawa N., 2000. Population field studies on the aphidophagous ladybird beetle Harmonia axyridis (Coleoptera: Coccinellidae): resource tracking and population characteristics. Popul. Ecol., 42(2), 115-127.
Saini E.D., 2004. Presencia de Harmonia axyridis (Pallas) (Coleoptera: coccinellidae) en la provincia de Buenos Aires. Aspectos Biologicos Morfologicos, 33, 151-160.
San Martin G., 2003. Étude de l'impact de l'urbanisation sur les populations de coccinelles à Bruxelles. Unpublished Licence en Biologie, Université Libre de Bruxelles (Belgique).
Snyder W.E. et al., 2004. Complementary biocontrol of aphids by the ladybird beetle Harmonia axyridis and the parasitoid Aphelinus asychis on greenhouse roses. Biol. Control, 30(2), 229-235.
Stuart R.J., Michaud J.P., Olsen L. & McCoy C.W., 2002. Lady beetles as potential predators of the root weevil Diaprepes abbreviatus (Coleoptera: Curculionidae) in Florida citrus. Florida Entomol., 85(3), 409-416.
Sun X.Q. et al., 1996. A preliminary study on the artificial diet of an aphidophagous coccinellid, Harmonia axyridis (Pallas) and its use to control strawberry aphids under plastic covering. J. Shanghai Agric. Coll., 14, 133-137.
Takahashi K. & Naito A., 1984. Seasonal occurrence of aphids and their predators (Col. Coccinellidae) in alfalfa fields. Bull. Natl Grassland Res. Inst., 29, 62-66.
Tedders W.L. & Schaefer P.W., 1994. Release and establishment of Harmonia axyridis (Coleoptera, Coccinellidae) in the Southeastern United-States. Entomol. News, 105(4), 228-243.
Thalji R., 2006. Composition of coccinellid communities in sugar beet fields in Vojvodina. Zbornik Matice Srpske za Prirodne Nauke, 2006(110), 267-273.
Tomov R., Trencheva K., Trenchev G. & Kenis M., 2009. The multicolored invasive Asian ladybird Harmonia axyridis (Pallas, 1773) (Coleoptera: Coccinellidae) new to the fauna of Bulgaria. Acta Zoologica Bulgarica, 61(3), 307-311.
Watanabe M., 2002. Cold tolerance and myo-inositol accumulation in overwintering adults of a lady beetle, Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol., 99(1), 5-9.
Wells M.L. & McPherson R.M., 1999. Population dynamics of three coccinellids in flue-cured tobacco and functional response of Hippodamia convergens (Coleoptera: Coccinellidae) feeding on tobacco aphids (Homoptera: Aphididae). Environ. Entomol., 28(4), 768-773.
Wells M.L., McPherson R.M., Ruberson J.R. & Herzog G.A., 2001. Coccinellids in cotton: population response to pesticide application and feeding response to cotton aphids (Homoptera: Aphididae). Environ. Entomol., 30(4), 785-793.
Wright E.J. & Laing J.E., 1980. Numerical response of coccinellids to aphids in corn in Southern Ontario. Can. Entomol., 112(10), 977-988.
Xue Y. et al., 2012. Intraguild predation of the aphid parasitoid Aphelinus certus by Coccinella septempunctata and Harmonia axyridis. BioControl, 57(5), 627-634.
Om dit artikel te citeren:
