
Diversité floristique sous canopée en plantation forestière de Mangombe-Edea (Cameroun)
Institut de la Recherche Agricole pour le Développement (IRAD). BP 219. CAM-Kribi (Cameroun). E-mail : njules_romain@hotmail.com – Université de Yaoundé I. Faculté des Sciences. Département de Biologie et Écologie végétale. BP 812. CAM-Yaoundé (Cameroun).
Université de Yaoundé I. Faculté des Sciences. Département de Biologie et Écologie végétale. BP 812. CAM-Yaoundé (Cameroun).
Université de Yaoundé I. Faculté des Sciences. Département de Biologie et Écologie végétale. BP 812. CAM-Yaoundé (Cameroun).
Museum national d’Histoire naturelle. 4 avenue du Petit Château. F-91800 Brunoy (France).
IRAD. Station de Bertoua. BP 203. CAM-Bertoua (Cameroun).
IRAD. Programme Forêt-Environnement. 2123 Kolbisson. CAM-Yaoundé (Cameroun).
Université de Yaoundé I. Faculté des Sciences. Département de Biologie et Écologie végétale. BP 812. CAM-Yaoundé (Cameroun).
Reçu le 12 septembre 2008, accepté le 11 septembre 2009
Résumé
La plantation d'arbres de Mangombé se trouve en forêt dense humide de basse altitude. Six parcelles dans ce site, dont une de Mansonia altissima (A.Chev.) A.Chev., deux de Lovoa trichilioides Harms, trois de Terminalia ivorensis A.Chev. et une forêt naturelle perturbée, ont fait l'objet d'une étude floristique. Le travail consiste en l'évaluation de la diversité floristique du sous-bois en vue d'apprécier l'incidence des plantations forestières sur la régénération naturelle. Au total, 26 familles, 42 genres et 46 espèces ont été recensés. Les Meliaceae et Apocynaceae forment le fonds floristique. Dans la parcelle 1 de T. ivorensis, M. altissima et la forêt naturelle, la faible valeur de l'équitabilité de Piélou traduit la dominance des effectifs par quelques espèces et la situation transitoire de l'écosystème. Par contre, le reste des parcelles présente une bonne répartition des individus au sein des espèces. On note une faible organisation du système écologique dans la parcelle de M. altissima, ce qui correspond à des conditions du milieu favorables à l'installation de nombreuses espèces représentées par un petit nombre d'individus. L'indice de Shannon relativement faible dans la parcelle 1 de T. ivorensis caractérise un peuplement où une espèce est dominante. Globalement, le sous-bois des parcelles se reconstitue progressivement et reste moins diversifié que la forêt naturelle environnante. La création d'un pool génétique au moyen d'entretiens sélectifs du sous-bois permettra une gestion polycyclique des parcelles dans le cadre de la production continue du bois.
Abstract
Floristic diversity under canopy in forest plantation of Mangombe-Edea (Cameroon). Trees plantation of Mangombe is situated in rain forest of low altitude. Six plots among which one of Mansonia altissima (A.Chev.) A.Chev., 2 of Lovoa trichilioides Harms, 3 of Terminalia ivorensis A.Chev. and one perturbed natural forest are concerned by this study. The work consisted to the evaluation of the floristic diversity under canopy in order to appreciate the influence of forest plantation on natural regeneration. A total of 26 families, 42 genders and 46 species were censured. Meliaceae and Apocynaceae are present in all the plots. In plot one of T. ivorensis, M. altissima and natural forest, the low value of equitability of Pielou traduces the dominance of flora by few species and the transitory situation of the ecosystem. On contrary the rest of plots present a good repartition of individuals among the species. One can notice a weak organization of the ecologic system in M. altissima plot, this corresponds to favourable conditions of environment for installation of many species represented by a few number of individuals. Shannon indices, relatively low in plot one of T. ivorensis characterize an ecosystem where one species is dominant. Globally, the regeneration under canopy is reconstituted progressively and remains less diversified than the nearest natural forest. Creation of genetic pool through selective entertainment under canopy will permit polycyclic management of plots for sustainable production of wood.
1. Plantation forestière et diversité dans le sous-bois
1Depuis quelques années, un intérêt croissant est porté aux forêts tropicales en raison de leur fonction économique dans les pays qui les possèdent, mais plus encore pour des raisons d’ordre écologique dans le changement global du climat et de conservation génétique (Grison, 1991). Au Cameroun, les écosystèmes forestiers couvrent 21 millions d’ha (Amine et al., 1992). Ils sont diversifiés, avec plus de 8 000 espèces de plantes parmi lesquelles plus de 300 espèces de bois exploitables (Letouzey, 1975). Les besoins croissants de l’homme à l’égard des forêts et le développement économique du pays essentiellement axé sur l’exploitation des ressources naturelles ont donné lieu à diverses formes d’exploitations qui ne garantissent pas toujours les capacités intrinsèques des ressources à régénérer (Poore et al., 1981 ; Singh, 1993 ; Chapman, 1995 ; Sonké, 1998). Ces pressions concurrentielles engendrent le recul de 100 000 à 200 000 ha de forêt par an, soit un taux annuel de déforestation de 0,3 à 1 % (Bikié et al., 2000 ; Kuyper et al., 2001). Ces pertes posent un problème de gestion et de conservation avec risque d’érosion génétique des espèces (Sonké, 1998). Face à ce contexte, Aubréville (1948) a recommandé au service forestier d’appuyer la régénération naturelle par les plantations forestières, sinon les forêts deviendraient industriellement inexploitables faute d’une densité suffisante de bois de qualité.
2À partir de 1930, le Cameroun s’est engagé dans une politique d’afforestation dont les objectifs variables (production du bois d’œuvre, de service et de feu, protection de l’environnement et amélioration des connaissances écologiques sur la dynamique forestière) sont compatibles avec les zones écologiques où ces plantations ont été installées. La plantation forestière de Mangombé, mise en place en 1964, a nécessité le défrichement de plus de 100 000 ha de forêt naturelle et moins de 500 ha ont été plantés. Elle visait la production de bois d’œuvre et la pérennisation du rôle des forêts au service des générations futures à travers l’amélioration des connaissances des fonctions écologiques et des dynamiques forestières (compositions, résilience, régénérations, interactions). Les difficultés à assurer l’entretien et le suivi de cette plantation forestière n’ont pas permis de tirer les connaissances nécessaires ; pourtant, en plus des difficultés techniques, sociales et économiques, les connaissances écologiques sur la dynamique forestière constituent encore un frein à l’élaboration des plans d’aménagement (Bertault et al., 1992). Globalement, les mécanismes régissant la dynamique des écosystèmes forestiers sont complexes, de l'établissement des plantules à la pollinisation et la dispersion des graines transportées par les animaux, l’eau ou le vent. Ces processus sont d'autant plus divers que l’écosystème abrite un large nombre d’espèces et de formes végétatives, comme c'est le cas dans les forêts tropicales humides (Chave, 2000). C’est ainsi que l’on se demande aujourd’hui : quel est l’impact des espèces plantées sur la diversité génétique du sous-bois ? Comment se présente la dynamique de régénération forestière ? Cette étude entend évaluer la diversité floristique sous la canopée en plantation forestière et vise l’amélioration des connaissances sur l’affirmation selon laquelle les plantations forestières monospécifiques contribuent à la simplification des écosystèmes.
2. Matériel et méthodes
3La plantation forestière de Mangombé (3°50’ N et 10°10’ E) est située à environ 16 km au Nord-Est de la ville d’Edéa en zone littorale camerounaise. La végétation originelle est une forêt ombrophile, dont les essences les plus fréquentes sont Lophira alata Banks ex C.F.Gaertn., Saccoglotis gabonensis (Baill.) Urb., Cynometra hankei Harms, Coula edulis Baill. Elle est riche en essence papetière avec un potentiel supérieur à 300 m3.ha-1 (Letouzey, 1968). La plantation a été progressivement mise en place à partir de 1964 par le Centre Technique Forestier Tropical (CTFT) à qui la recherche forestière de la République du Cameroun a été confiée après les indépendances et reprise en 1975 par l’Office National de Régénération des Forêts (ONAREF), puis par l’Institut de la Recherche Agricole pour le Développement (IRAD). La plantation est située en plaine côtière, à environ 30 m d’altitude. Les rivières et eaux de ruissellement ont quelque peu érodé le site et contribué à donner au relief une morphologie collinaire de pentes faibles estimées à 25 %. La croissance démographique dans les villages riverains des plantations a accru la demande en ressources foncières et produits forestiers, ce qui engendre des pressions diverses à la périphérie des plantations, notamment l’extension des parcelles agricoles dont le système est itinérant sur brûlis, l’urbanisation et la récolte des produits forestiers.
2.1. Dispositif expérimental
4Pour l’inventaire de la régénération naturelle dans le sous-bois, trois à quatre placettes de 10 x 10 m, soit 300 à 400 m2, ont été installées dans chaque parcelle de superficie unitaire de 0,3 à 0,5 ha. L’installation des placettes prend en compte la superficie des parcelles et l’hétérogénéité du sous-bois (Loumeto et al., 1997). Au total, six parcelles plantées et une parcelle installée en forêt naturelle environnant la plantation ont fait l’objet d’étude floristique. Il s’agit de : une parcelle de Mansonia altissima (A.Chev.) A.Chev. (Bété), série 1989 ; deux parcelles de Lovoa trichilioides Harms (Bibolo), donc 872 A et 872 B, série 1987 ; trois parcelles de Terminalia ivorensis A.Chev. (Framiré), série 1983.
5L’absence de banques de données sur la plantation constitue une des véritables contraintes rencontrées lors de l’exécution de ce travail.
2.2. Inventaire floristique des recrûs
6À l’intérieur des placettes, tous les recrûs d’au moins 5 cm de diamètre ont été marqués, recensés, identifiés et leurs circonférences mesurées. Les échantillons non identifiés sur le terrain l’ont été à l’Herbier National du Cameroun.
7Plusieurs indices ont été utilisés pour mettre en évidence la diversité spécifique des communautés végétales :
8– l’indice de diversité de Shannon (1949) cité par Guedje (2002) :
9ISH = - ∑ (Ni/N) log2 (Ni/N) s’exprime en bit
10où Ni = effectif de l’espèce i ; N = effectif total des espèces,
11– l’équitabilité de Pielou (1966) cité par Tchoumi (2001) est le rapport de la diversité d’un peuplement ou d’un échantillon et le nombre N d’espèces présentes dans la parcelle. Il exprime la régularité, la répartition équitable des individus au sein des espèces :
12EQ = ISH/log2 (N)
13– la richesse spécifique RS est le nombre total d’espèces de la distribution observée,
14– l’indice de diversité de Simpson (Colinvaux, 19861 cité par Tchoumi, 2001) représente la probabilité pour que deux individus pris au hasard dans le peuplement étudié appartiennent à la même espèce. Il mesure la manière avec laquelle les individus se repartissent entre les espèces d’une communauté :
15D’ = ∑ (Ni/N)2
16– la diversité des familles est l’expression en pourcentage d’apparition de chacune des familles inventoriées dans le sous-bois,
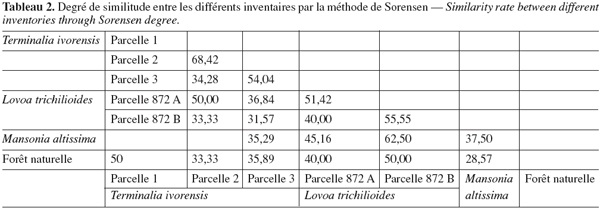
17– le coefficient de similarité de Sorensen (1948), cité par Sonke (1998), permet de savoir si deux parcelles a et b comparées sur le plan floristique appartiennent à la même communauté végétale. Il s’agit du pourcentage d’espèces communes dans deux parcelles. Pour une valeur de K supérieure à 50 %, on peut conclure que les deux parcelles comparées appartiennent à une même communauté végétale. Dans le cas contraire, les parcelles appartiennent à des communautés végétales différentes :
18K = (2C/A+B) x 100
19A = nombre d’espèces de la parcelle a
20B = nombre d’espèces de la parcelle b
21C = nombre total des espèces communes aux parcelles a et b
3. Résultats
3.1. Composition floristique : diversité spécifique, générique et abondance des familles
22Parcelles de Terminalia ivorensis (Framiré). La plantation 833 de T. ivorensis comporte trois parcelles (1, 2 et 3), ayant chacune 0,34 ha de superficie. Ces parcelles ont fait l’objet d’un essai d’écartements de valeurs respectives : 3,5 x 3 m ; 3,5 x 4 m et 3,5 x 3,5 m.
23L’abondance des espèces varie avec la méthode sylvicole. Les effectifs vont décroissant avec l’augmentation des écartements. Dans la parcelle 1 où les écartements sont inférieurs à ceux des autres parcelles, 75 individus (2 500 tiges par ha) sont enregistrés. Cet effectif est constitué de 18 espèces réparties en 17 genres et 15 familles. Trois espèces représentent 34,6 % des tiges, il s’agit de Macaranga hurifolia Beille (14,6 %), Canarium schweinfurthii Engl. (12 %) et Rauvolfia vomitoria Afzel. (8 %).
24La parcelle 2 compte 15 familles comprenant 18 genres et 20 espèces pour un total de 66 individus (2 200 tiges par ha), tandis que la parcelle 3 possède l’effectif le plus réduit, 44 individus où 4 espèces, Greenwayodendron suaveolens (Engl. & Diels) Verdc. (11,4 %), Guibourtia demeusei (Harms) Leonard (11,4 %), Carpolobia lutea G.Don (13,6 %) et L. trichilioides (13,6 %) représentent 50 % des effectifs.
25Au total, 5 familles, Passifloraceae, Euphorbiaceae, Annonaceae, Apocynaceae, Ochnaceae et 6 espèces, R. vomitoria, G. suaveolens, C. schweinfurthii, L. alata, L. trichilioides et Barteria fistulosa (Mast.) Sleumer sont communes à toutes les parcelles.
26L’indice de diversité de Shannon (ISH) est compris entre 1,19 et 2,70 bits pour l’ensemble des parcelles. L’équitabilité de Piélou varie entre 0,27 et 0,69. L’indice de diversité de Simpson (D’) 0,075-0,086 varie peu entre les trois parcelles. Ces différents indices présentent des variations directement liées aux nombres d’espèces et d’individus inventoriés et à la répartition des individus entre les espèces. La richesse spécifique entre les parcelles varie peu, de même que la probabilité pour que deux individus pris au hasard dans les peuplements appartiennent à la même espèce.
27Parcelles de Lovoa trichilioides (Bibolo). Les parcelles 872 A et 872 B portent sur des essais d’écartements respectifs de 3 x 3 m et 4 x 4 m. Au total, 44 individus, soit 1 466 tiges par ha (parcelle 872 B) et 56 individus, soit 1 866 tiges par ha (parcelle 872 A), ont été inventoriés.
28La richesse spécifique du sous-bois est identique dans les deux parcelles qui comptent chacune 18 espèces. Ces espèces sont reparties en 17 genres et 13 familles dans la parcelle 872 B, la parcelle 872 A compte 17 genres et 12 familles.
29Dans la parcelle 872 A, les Caesalpiniaceae et Apocynaceae constituées de 3 espèces chacune, soit une proportion de 16,6 % du nombre d’espèces de la flore, sont les groupes floristiques les plus diversifiés. Les Euphorbiaceae comptent 3 espèces pour une proportion de 16,6 % du nombre d’espèces de la flore, c’est le groupe floristique le plus diversifié dans la parcelle 872 B. Cinq espèces présentent une abondance remarquable dans la parcelle 872 B, il s’agit de C. lutea, Diospyros bipindensis Gürke, C. schweinfurthii, Diospyros crassiflora H.Perrier et Khaya ivorensis A.Chev. estimées à 45,5 % de la flore.
30Le sous-bois de la parcelle 872 A possède 2 espèces dominantes, R. vomitoria et D. crassiflora dont le total cumulé d’individus représente 28,5 % de la flore.
31Au total 9 espèces, Alstonia boonei De Wild., M. hurifolia, R. vomitoria, D. crassiflora, C. schweinfurthii, Desplatsia dewevrei (De Wild. & T.Durand) Burret, Syzygium guineense (Willd.), Strombosia pustulata Oliv. et D. bipindensis sont communes aux deux parcelles.
32L’indice de diversité de Shannon (ISH) est de 2,68 bits (parcelle 872 A) et 2,74 bits (parcelle 872 B). L’équitabilité de Pielou varie entre 0,66 et 0,72. L’indice de diversité de Simpson 0,080 et 0,073 varie faiblement. En général, la probabilité pour que deux individus pris au hasard appartiennent à la même espèce est faible dans les deux parcelles, même si elle tend à être un peu plus élevée dans la parcelle 872 A.
33Parcelle de Mansonia altissima (Bété). Cette parcelle couvre 0,44 ha et a fait l’objet d’un essai d’écartement de 4 x 4 m. Nonante-cinq individus, soit 3 166 tiges par ha, ont été recensés dans le sous-bois.
34L’inventaire floristique au sein de cette parcelle a permis de recenser 9 familles comprenant 12 genres et 13 espèces. Les Apocynaceae représentent le groupe floristique le plus diversifié avec 3 espèces, Voacanga africana Stapf, R. vomitoria et A. boonei. Cette famille compte 80 individus qui représentent 84,21 % de l’effectif total.
35Les Sapotaceae et Myristicaceae sont les groupes floristiques les moins diversifiés du peuplement. Chacune de ces familles compte une seule espèce et représente 1,05 % de l’effectif total du peuplement. En général, V. africana présente un effectif remarquable (64,21 %), alors que pour le reste du peuplement l’abondance varie peu d’une espèce à l’autre.
36La variation de quelques paramètres du peuplement montre que la probabilité pour que deux individus pris au hasard dans le peuplement appartiennent à la même espèce est élevée, elle est estimée à 0,42. L’indice de diversité de Shannon a pour valeur 1,49 bits et l’équitabilité de Pielou calculée pour ce peuplement forestier est de 0,33.
3.2. Caractérisation du point de vue floristique de la forêt naturelle
37L’inventaire a permis de recenser en forêt naturelle 119 individus, soit 2 975 tiges par ha. La flore du sous-bois comprend 22 espèces réparties en 22 genres et 15 familles. La famille des Euphorbiaceae qui compte les 4 espèces, Mareyopsis longifolia (Pax) Pax & K.Hoffm., Drypetes gossweileri S.Moore, M. hurifolia, Ricinodendron heudelotii (Baill.) Pierre ex Heckel, est le groupe floristique le plus diversifié. Ce groupe représente 7,56 % de l’effectif de la flore. Les Apocynaceae comprennent deux espèces, V. africana (37,81 %) et R. vomitoria (5,03 %). Cette famille compte 51 individus, soit 42,85 % des effectifs.
38V. africana et S. pustulata sont par ordre d’importance décroissante les espèces les plus représentées avec une proportion respective de 37,81 % et 8,40 % de la flore.
39La richesse spécifique du sous-bois en forêt naturelle est importante comparée à celle des peuplements artificiels. La probabilité pour que deux individus pris au hasard dans le peuplement étudié appartiennent à la même espèce est relativement élevée, l’indice de diversité de Simpson est estimé à 0,17, l’équitabilité de Pielou 0,50 et l’indice de diversité de Shannon 2,4. (Tableau 1).
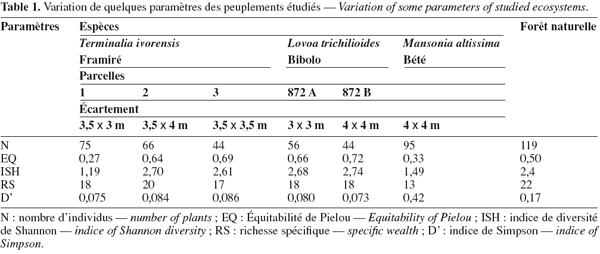
3.3. Fonds floristique des peuplements étudiés
40Au total, 499 individus ont été recensés dans l’ensemble des parcelles retenues dans le cadre de cette étude. Ils sont repartis dans 26 familles représentées par 42 genres et 46 espèces.
41Les Meliaceae et Apocynaceae forment le fonds floristique, ces deux groupes floristiques sont présents dans tous les peuplements. Cependant, une seule espèce, R. vomitoria, est commune à toutes les parcelles. Cette espèce représente 7 % de la flore inventoriée. Les Apocynaceae, estimées à 32,06 % de la flore, sont le groupe floristique le plus important.
42Quelques familles sont très faiblement représentées, il s’agit des Rhizophoraceae (0,8 %), Myristicaceae (1 %) et Rhamnaceae (0,6 %) dont l’effectif cumulé représente 2,4 % de la flore de l’ensemble des parcelles. La famille des Rhamnaceae n’est propre qu’à la parcelle 1 de T. ivorensis.
43Une comparaison de la composition spécifique des différents inventaires indique un seuil de similarité qui varie de 28,57 à 68,42 %. Ces fortes valeurs du coefficient de Sorensen traduisent une grande homogénéité entre les parcelles 1 et 2 de T. ivorensis ; de L. trichilioides (872 A) et T. ivorensis (parcelle 1) ; de T. ivorensis (parcelles 3 et 2), de T. ivorensis (parcelle 3) et L. trichilioides (872 A) ; de L. trichilioides (872 B et 872 A) ; de L. trichilioides (872 A) et M. altissima ; de L. trichilioides (872 B) et la forêt naturelle ; de la forêt naturelle et T. ivorensis (parcelle 1). Ces parcelles appartiennent à une même communauté végétale (Tableau 2).
4. Discussion
4.1. Comportement du sous-bois des plantations forestières
44La régénération naturelle est un ensemencement spontané qui permet de comprendre la dynamique forestière marquée par l’existence de trouées et le rôle de la composition « floristique initiale » ou potentiel floristique (Alexandre, 1989). La dynamique de population quant à elle désigne l’ensemble des changements qui se produisent au cours de la vie d’une population. Cet ensemble comprend le recrutement et la croissance, la sénilité, la mortalité, les fluctuations saisonnières de la biomasse, la stabilité de chaque classe d’âge et son importance relative, plus les effets que l’un ou plusieurs de ces facteurs exercent sur la population. Pour Blondel (1979), c’est une succession écologique, c'est-à-dire un ensemble de processus par lesquels un écosystème naturellement ou artificiellement altéré ou détruit, entreprend spontanément de se reconstituer pour recouvrer un état qui soit en quelque sorte similaire à l’état initial. Ces conceptions de la dynamique semblent bien indiquées pour cette étude.
45La reconstitution du sous-bois forestier varie d’un peuplement à l’autre. Elle est importante dans la parcelle de M. altissima (3 166 tiges par ha) suivi de la forêt naturelle (2 975 tiges par ha). Cette forte inégalité de la régénération forestière dans le sous-bois peut s’expliquer par la différence entre les années de mise en place des parcelles et les conditions écologiques favorables. Au sein d’une même série, c'est-à-dire les parcelles créées la même année, nous pouvons noter une forte colonisation des trouées forestières suite à l’ouverture de la canopée. Ndotar (2004) et Hubbell et al. (1999) confirment que le recrutement dans les chablis est plus important que celui rencontré en forêt fermée. Ce phénomène, appelé chablis, est considéré comme un des moteurs de la sylvigenèse car il favorise l'arrivée de la lumière au sol et donc la régénération de nouveaux individus (Riéra, 1983). L’étude de la succession végétale dans les abords de l’axe routier Yaoundé – Douala (Cameroun) montre que la recolonisation végétale s’est faite avec les stades successifs (Mapongmetsem, 1987) :
46– la phase herbacée constituée d’espèces rudérales dont la taille avoisine 30 cm. Elle regroupe Euphorbia heterophylla L., Synedrella nodiflora (L.) Gaertn., Aspilia africana (P.Beauv.) C.D.Adams, Bidens pilosa L., Erigeron floribundus (Kunth) Sch.Bip., Chromolaena odorata (L.) King & Rob. Ces espèces seront par la suite envahies par les graminées comme Setaria megaphylla (Steud.) T.Durand & Schinz, Pennisetum purpureum Schumach., Hyparrhenia diplandra (Hack.) Stapf ;
47– la phase arbustive marquée par l’occupation du tapis graminéen par les espèces ligneuses de faible taille, estimée à 50 cm. Elle comprend Musanga cecropioides R.Br., Anthocleista schweinfurthii Gilg, Trema orientalis Blume, Ceiba pentandra (L.) Gaertn., Harungana madagascariensis Lam. ex Poir., Albizia adianthifolia (Schumach.) W.Wight, Alchornea cordifolia Müll.Arg. ;
48– la phase arborée constituée des essences forestières à bois dur, composée de Alstonia boonei, Lophira alata, Piptadeniastrum africanum (Hook.f.) Brenan, Terminalia superba Engl. & Diels, Desbordesia glaucescens (Engl.) Tiegh., Milicia excelsa (Welw.) C.C.Berg.
4.2. Diversité des familles dans les parcelles
49Les peuplements sondés comptent 26 familles, alors que les études de Ndotar (2004) révèlent la présence de 35 familles dans les plantations d’essences papetières contigües à notre site d’étude. Les peuplements se caractérisent par une variation dans la diversité des familles. Certaines familles sont présentes dans toutes les parcelles ; il s’agit des Euphorbiaceae, Apocynaceae, Meliaceae, Caeasalpiniaceae ; d’autres, par contre, appartiennent exclusivement à une seule parcelle, ce sont : Rhamnaceae et Verbenaceae (parcelle 1 à T. ivorensis), Bombacaceae (parcelle 2 à T. ivorensis), Rubiaceae (parcelle 3 à T. ivorensis), Moraceae (parcelle 872 B à L. trichilioides), Fabaceae (M. altissima), Cecropiaceae (forêt naturelle). Cette étude a identifié six nouvelles familles : Clusiaceae, Passifloraceae, Rhizophoraceae, Polygalaceae, Myrtaceae et Fabaceae, en dehors de celles obtenues par Ndotar (2004) ; ceci porte la diversité floristique du sous-bois des plantations de Mangombé à 41 familles.
4.3. Diversité spécifique des parcelles sondées
50Les écosystèmes matures n’ayant pas connu des perturbations ont une équitabilité optimale de l’ordre de 0,6 à 0,8 (Odum, 19762 cité par Sonke, 1998) ; c’est le cas des parcelles étudiées, à l’exception de celles de T. ivorensis (parcelle 1) (0,27), M. altissima (0,33) et la forêt naturelle (0,50) qui enregistrent de petites valeurs d’équitabilité. Ces faibles valeurs traduisent la dominance des effectifs par quelques espèces, elles peuvent aussi indiquer l’état de stress ou la situation transitoire de l’écosystème.
51Les parcelles 3 et 2 de T. ivorensis et de L. trichilioides 872 B ont des valeurs élevées d’équitabilité, respectivement estimées à 0,69 ; 0,64 et 0,72. Ces valeurs traduisent une bonne répartition des individus au sein des espèces.
52L’indice de Shannon varie d’un peuplement à l’autre. La valeur la plus élevée, 2,74 bits, est obtenue dans la parcelle de L. trichilioides (872 B), elle est inférieure à la moyenne 4,95-4,80 bits dans une parcelle similaire de Bilik (Pouentoumoun, 2005). Cette forte valeur traduit une grande diversité et une bonne reconstitution de la diversité floristique du sous-bois, sans doute en raison des conditions favorables du milieu. Pour l’ensemble des peuplements étudiés, les valeurs de l’indice de Shannon trouvées sont faibles par rapport à celles obtenues en forêts atlantiques biafréennnes à Caesalpiniaceae (Guedje, 2002) et dans la forêt de la réserve de la Biosphère du Dja, à l’exception de la forêt à Gilbertiodendron dewevrei (De Wild.) J.Léonard (ISH = 2,7) (Sonke, 1998).
53L’indice de Simpson est très sensible à la répartition des individus entre les espèces. Les fortes valeurs de cet indice expriment une faible organisation du système écologique et correspondent selon Dajoz (1982) à des conditions du milieu favorables à l’installation de nombreuses espèces représentées par un petit nombre d’individus, alors que les faibles valeurs de l’indice de Shannon sont caractéristiques des peuplements où une espèce est dominante. En Guyane, Sabatier (1990) et Lang et al. (1983) remarquent une structure typique des forêts tropicales dans une réserve, avec un nombre important d'espèces, représentées par un faible effectif.
54Les perturbations naturelles, chablis et volis, et la dissémination des graines et pollens dans les parcelles étudiées provoquent des variations dans le stade successionnel de la forêt et, par conséquent, des fluctuations dans les proportions des différentes espèces. La présence de trouées dans la canopée joue un rôle fondamental, elle se caractérise par un stade pionnier où les individus adultes qui vont composer le peuplement sont des espèces pionnières ou héliophiles, susceptibles de ne germer que dans de bonnes conditions de lumière. Ce groupe d’essences se distingue par une croissance très rapide, une maturité précoce, une fécondité élevée et une espérance de vie faible. Des résultats similaires sont obtenus à la suite de l’étude des caractéristiques écologiques intrinsèques des taxons dans la réserve naturelle des Nouragues en Guyane française (Riera, 1983 ; Riera et al., 1990 ; Jean, 2001 ; Trouillon, 2006). L’augmentation du nombre de trouées dans les parcelles va favoriser l’installation d’espèces pionnières. Ces taxons qui nécessitent de la lumière pour leur germination croissent rapidement, leurs floraisons et fructifications sont précoces et abondantes mais leur espérance de vie relativement courte (Trouillon, 2006). Dans une forêt mature par contre, on observe plus d'espèces sciaphiles ou tolérantes à l'ombre. Ces dernières ont besoin de lumière pendant les stades juvéniles de leur développement avant d'atteindre la canopée et se caractérisent par une maturité tardive, une fécondité et une densité faibles ainsi qu'une espérance de vie très importante. Ganglo (2005), dans une étude sur les groupements de sous-bois réalisée dans le périmètre du bois de feu de la Lama au Bénin, a identifié quatre groupements végétaux ayant une richesse spécifique moyenne par relevé estimée à environ 26-39 espèces. L’indice de Shannon dans ce bois est compris entre 1,89 et 2,40, alors que le coefficient d’équitabilité de Pielou est de 0,39 à 0,49. La station la plus productive indiquée par le groupement végétal à Deinbollia pinnata Schum. & Thonn. parait propice à des plantations de bois d’œuvre de qualité à partir d’espèces comme Afzelia africana Sm., Pterocarpus erinaceus Poir., Antiaris toxicaria (Pers.) Lesch., Milicia excelsa, Sterculia tragacantha Lindl.
4.4. Abondance spécifique
55Présenter la structure complexe d’une communauté uniquement par sa richesse spécifique ou un indice de diversité donne des informations incomplètes. L’étude statistique de l’abondance des espèces d’une communauté végétale présente une grande importance. En effet, elle permet de mieux interpréter la nature des interactions entre espèces et de mettre en évidence les facteurs qui déterminent leur fréquence relative.
56Au total, 42 genres et 46 espèces ont été inventoriés. La richesse spécifique est plus élevée en forêt naturelle (22 espèces), cette valeur est inférieure à 199 espèces et 65 espèces respectivement obtenues par Guedje et al. (2002) dans les forêts à Garcinia lucida Vesque du sud Cameroun et Pouentoumoun (2005) dans les plantations forestières de Bilik en forêt semi-caducifoliée. Certains auteurs soutiennent que la diversité spécifique des groupements végétaux est très élevée durant les premiers stades de la succession avec une réduction du nombre d’individus vers le stade climacique ; pour d’autres, la structure du peuplement se met en place dès les premières années de succession et la maturation forestière n’entraine qu’une baisse notable de la richesse spécifique (Sonke, 1998). Sonke (1998) souligne que la tendance à la croissance de la richesse spécifique, la densité et la diversité des peuplements constitue une loi générale pour toutes les successions écologiques.
57La forte richesse spécifique de certains peuplements peut s’expliquer par les conditions écologiques qui seraient favorables à la régénération des espèces présentes. On note une faible richesse spécifique dans la parcelle de M. altissima. Cette situation pourrait s’expliquer par la densité de la canopée des peuplements environnants susceptibles de constituer un frein à la dissémination des diaspores responsables du recrutement. La pente du relief serait aussi favorable au ruissellement des eaux avec pour conséquence une rapide variation du gradient de fertilité.
58Plusieurs familles se caractérisent par leur abondance en tiges, il s'agit par ordre d’importance décroissante d'Apocynaceae (32,26 %), Ebenaceae (6,8 %), Burseraceae (6,4 %), Meliaceae (6,2 %) et Euphorbiaceae (5,8 %). On peut remarquer la présence des Cecropiaceae représentés par Musanga cecropioides, c’est une espèce qui colonise les forêts secondaires et trouées forestières ; elle est aussi liée à la présence de l’homme, étant donné que le site de Mangombé fut autrefois habité par les populations qui se sont déportées lors de la création des plantations vers l’ancienne route Douala – Edéa.
59Pour enrichir la forêt, le rôle du sylviculteur consiste à favoriser le développement des essences utiles au détriment des autres. Deux méthodes sont possibles, relevant chacune d’une conception différente :
60– on peut considérer que le milieu naturel n’a guère de valeur que par son sol et son climat et que la végétation existante n’y est pas capable d’évoluer vers un état économiquement rentable et en tout cas, les efforts du sylviculteur en ce sens seraient sans rapport avec le résultat qu’il pourrait atteindre. On s’oriente alors vers une méthode d’enrichissement par plantation dans les layons ouverts en forêt ou la plantation en parcelle,
61– on peut au contraire estimer que les différents peuplements dans la forêt secondaire représentent, sous forme de semis, gaules, perches, arbres petits ou moyens d’espèces utiles, un potentiel de richesse en bois d’œuvre qui n’est pas à négliger. On envisage alors une méthode d’enrichissement basée sur la végétation existante et sur sa régénération naturelle. Sur cette dernière conception a été fondée la méthode d’enrichissement mise en œuvre depuis 1947 au Cameroun dans la forêt domaniale de Mbalmayo. Les comptages effectués en 1950 sur 1 570 ha montrent un taux de survie de 48,5 % sur un total initial de 105 359 tiges plantées en layons (Morellet, 1952). On note également que 23 866 semis issus de régénération naturelle et appartenant à une douzaine d’espèces utiles avaient pris place sur ces layons inventoriés. Ceci relève à 75,5 % le pourcentage des plants existant par rapport à ceux plantés primitivement. Il est à remarquer que la proportion de semis naturel est certainement plus forte. Les semis naturels existants entre les layons n’ont pas été dénombrés. Cette régénération naturelle accessoire a donc constitué pour les plantations en layons un appoint précieux. Dans certaines parcelles, le nombre de plants issus de semis naturel est même supérieur au nombre des arbres plantés (Morellet, 1952).
5. Plantation forestière de Mangombé entre le présent et l'avenir
62Actuellement, quelques points d’ombres persistent, cela est sûrement dû au manque d’archives et d’informations sur l’historique des parcelles. La présente étude révèle une biodiversité végétale importante dans les parcelles étudiées. Elle se caractérise par une diversité spécifique, générique et des familles. L’étude floristique des recrûs a permis de recenser 42 genres, 46 espèces et 26 familles. La biodiversité au sein d'une parcelle est fonction d'un pool d'espèces potentiellement disponibles et des conditions permettant leur établissement. L'hétérogénéité observée dans la distribution spatiale des espèces épouse le concept de mosaïque défini par Aubréville (1938).
63Six nouvelles familles regroupant les Clusiaceae, Passifloraceae, Rhizophoraceae, Polygalaceae, Myrtaceae et Fabaceae ont été inventoriées en dehors des 35 familles obtenues par Ndokar (2004), ce qui porte à 41 familles la diversité du sous-bois des plantations de Mangombé.
64Les plantations forestières étudiées, de par la valeur de leur équitabilité (0,6-0,8), sont assimilables à des écosystèmes peu perturbés, à l’exception des parcelles de T. ivorensis (parcelle 1), M. altissima et la forêt naturelle qui enregistrent de petites valeurs d’équitabilité (EQ < 0,5). Ces faibles valeurs traduisent la dominance des effectifs par quelques espèces ou la situation transitoire de l’écosystème.
65Les Meliaceae et Apocynaceae forment le fonds floristique. Une seule espèce, R. vomitoria, est commune à toutes les parcelles. Cette espèce représente 7 % de l’effectif total de la flore inventoriée. Les Apocynaceae sont le groupe floristique le plus important avec une proportion de 32,06 % de la flore.
66Les parcelles étudiées sont des mosaïques d’unités dans des états dynamiques différents, donc la dynamique de régénération naturelle dépend de la nature et de l’intensité des perturbations naturelles (chablis, volis), y compris des mécanismes de dissémination des diaspores. Ces modifications de structure entretiennent la dynamique et la diversité spécifique des écosystèmes. En général, la régénération naturelle dans le sous-bois n’est pas négligeable, les paramètres mesurés indiquent une reconstitution progressive de la flore. C’est ainsi que les plantations forestières peuvent, par leur diversité et leur souplesse de mise en œuvre, contribuer efficacement à satisfaire les besoins en produits forestiers et participer au développement économique des pays. Dans leur diversité, les plantations sont un des outils au service de l'objectif d'aménagement durable des forêts des régions tropicales (Marien et al., 2004). Au Gabon, Doucet et al. (2004) ont obtenu des résultats très encourageants et proposent des techniques relativement simples et peu onéreuses pour assister la régénération naturelle de l’Okoumé. Même si Foury (1956) reconnait que la régénération naturelle peut donner techniquement des résultats satisfaisants, notamment avec certaines essences envahissantes et dans certaines conditions du milieu, cet auteur préfère cependant la régénération artificielle qu’il estime comme étant une méthode techniquement sûre et plus rentable dans l’avenir.
67Étant donné l’importance de la régénération dans le sous-bois des plantations étudiées, il serait intéressant d’envisager une exploitation polycyclique dans l’optique de pérenniser la production de bois. Pour ce faire, il convient de :
68– pratiquer les entretiens sélectifs du sous-bois qui épargnent les recrûs bien conformés d’essences commerciales, en vue de créer un pool génétique varié et plus large,
69– exploiter les arbres ayant atteint le diamètre moyen d’exploitation, ceci réduit la concurrence avec les espèces de l’étage inférieur.
Bibliographie
Alexandre D.Y., 1989. Dynamique de la régénération naturelle en forêt dense de la Côte d’Ivoire. Paris : ORSTOM.
Amine M. & Bessong J.B., 1992. Conserving biological diversity in managed tropical forest. In: Blockhus J.M., Dillenbeck M. & Sayer J.A. The IUCN Forest conservation programme. Gland, Switzerland: IUCN; Yokohama, Japan: ITTO, 1-15.
Aubréville A., 1938. La forêt coloniale. Les forêts de l’Afrique occidentale française. Ann. Acad. Sci. Colon., 9, 1-245.
Aubréville A., 1948. Richesse et misère des forêts de l’Afrique noire française. In : Mission forestière AEF - Cameroun - AOF 1945-1946. Paris : Jouve, 160-170.
Bertault J.G., Dupuy B. & Maitre H.F., 1998. Recherches sylvicoles pour un aménagement durable en forêt dense humide. In : Actes du Congrès IUFRO, Berlin, Allemagne, 5-23.
Bikié H., Ndoye O. & Sunderlin W.D., 2000. L’impact de la crise économique sur les systèmes agricoles et le changement du couvert forestier dans la zone forestière humide du Cameroun. Yaoundé : Center for International Forestry Research (CIFOR), 1-40.
Blondel J., 1979. Biogéographie et écologie. Paris : Masson.
Chapman A.C., 1995. Primate seed dispersal. Coevolution and conservation implications. In: Bawa K.S. & Hadley M., eds. Reproductive ecology of tropical forests plants. Paris : UNESCO, 30-50.
Chave J., 2000. Dynamique spatio-temporelle de la forêt tropicale. Ann. Phys. Fr., 25(6), 1-184.
Dajoz R., 1982. Précis d’écologie. 4e éd. Paris : Bordas.
Doucet J.-L., Ntchandi Otimbo P.-A. & Boubady A.-G., 2004. Comment assister la régénération naturelle de l'Okoumé dans les concessions forestières ? Bois Forêts Trop., 279(1), 59-71.
Foury P., 1956. Comparaison des méthodes d’enrichissement utilisées en forêt dense équatoriale. Bois Forêts Trop., 47, 14-25.
Ganglo J.C., 2005. Groupement de sous-bois, identification et caractérisation des stations forestières : cas d’un bois au Bénin. Bois Forêts Trop., 285(3), 35-46.
Grison F., 1991. Recherche forestière internationale. Le temps des grandes manœuvres pour de nouvelles ambitions. Bois Forêts Trop., 227, 75-83.
Guedje M.N., 2002. La gestion des populations d’arbres comme outil pour une exploitation durable des produits forestiers non ligneux : l’exemple de Garcia lucida (Sud Cameroun). Thèse de doctorat : Université Libre de Bruxelles (Belgique).
Guedje M.N., Nkongmeneck B.A. & Lejoly J., 2002. Composition floristique et structure des formations à Garcinia lucida dans la region de Bipindi, Akom II (Sud Cameroun). Acta Bot. Gallica, 149(2), 157-178.
Hubbell S.P. et al., 1999. Light gap disturbance, recruitment, limitation and tree diversity in a neotropical forest. Science, 283(5401), 554-557.
Jean O., 2001. Perturbations naturelles, biomasse et dynamique en forêt tropicale humide en Guyane. Mémoire de DEA : Université Paul Sabatier, Toulouse III (France).
Kuyper T.W., Onguene N.A., Van Gemerden B.S. & Mbelli H., 2001. Integration of ecological knowledge in sustainable management of rainforests of south Cameroon, with special reference to myorrhyzia associations. Wageningen, The Netherlands: The Tropenbos Foundation, 5-27.
Lang G.E. & Knight D.H., 1983. Tree growth, mortality, recruitment and canopy gap formation during a ten years period in a tropical moist forest. Ecology, 64(5), 1075-1080.
Letouzey R., 1968. Étude phytogéographique du Cameroun. Paris : Édition Lechevalier.
Letouzey R., 1975. Notice de la carte phytogéographique du Cameroun au 1/500 000. Domaine de la forêt dense humide toujours verte. Toulouse, France : Institut de la Carte Internationale de la Végétation ; Yaoundé : Institut de la Recherche Agronomique (Herbier National), 62-142.
Loumeto J.J. & Bernhard-Reversad F., 1997. La biodiversité dans les plantations d’arbres à croissance rapide au Congo. Bois Forêts Trop., 253(3), 57-61.
Mapongmetsem P.M., 1987. Étude de la recolonisation végétale des abords de l’axe lourd Yaoundé – Douala (zone comprise entre M’van et Mbankomo). Mémoire de maitrise : Université de Yaoundé (Cameroun).
Marien J.N. & Mallet B., 2004. Nouvelles perspectives pour les plantations forestières en Afrique centrale. Bois Forêts Trop., 282, 67-79.
Morellet J., 1952. Une expérience sylvicole au Cameroun. Bois Forêts Trop., 25, 297-312.
Ndotar M., 2004. Diversité floristique des recrûs dans les plantations de la Cellucam près d’Edea. Mémoire de DEA : Université de Yaoundé I (Cameroun).
Poore D. & Sayer J., 1981. The management of tropical moist forest land: ecological guidelines. 2nd ed. Gland, Switzerland; Cambridge, UK: IUCN.
Pouentoumoun J.M., 2005. Biodiversité floristique dans les plantations : cas des peuplements forestiers plantés de Bilik (Mbalmayo). Mémoire de DEA : Université de Yaoundé I (Cameroun).
Riera B., 1983. Chablis et cicatrisation en forêt guyannaise (Piste de Saint Elis). Thèse de doctorat : Université Paul Sabatier, Toulouse (France).
Riera B., Puig H. & Lescure J.P., 1990. La dynamique de la forêt naturelle. Bois Forêts Trop., spécial Guyanne, 219, 69-78.
Sabatier D., 1990. Gestion de l’écosystème forestier et aménagement de l’espace régional. Actes du 2e Congrès régional de l'environnement, 16-17 février 1990, Cayenne, Guyane.
Singh K.D., 1993. L'évaluation des ressources forestières tropicales. Unasylva, 44(174), 10-20.
Sonke B., 1998. Études floristiques et structurales des forêts de la réserve de faune du Dja (Cameroun). Thèse de doctorat : Université Libre de Bruxelles (Belgique).
Tchoumi N.F., 2001. Contribution à l’étude écologique et structurale des forêts somitales du massif du Mbam Minkoum (région de Yaoundé). Mémoire de maitrise : Université de Yaoundé I (Cameroun).
Trouillon A., 2006. Étude de la dynamique forestière en Guyane française en relation avec les traits d'histoire de vie et les perturbations naturelles. Mémoire de master 2 : Université Pierre et Marie Curie, Paris (France).
Notes
Pour citer cet article
