
Influence du type de fertilisation, du jour de pâturage et du mois de l'année sur les rejets azotés des vaches laitières
Univ. Liège. Faculté de Médecine vétérinaire. Service de Nutrition. Chemin de la Ferme, 6/39. B-4000 Liège (Belgique). E-mail : Isabelle.dufrasne@ulg.ac.be
Univ. Liège. Faculté de Médecine vétérinaire. Service de Nutrition. Boulevard de Colonster, 20/43. B-4000 Liège (Belgique).
Univ. Liège. Faculté de Médecine vétérinaire. Service de Nutrition. Boulevard de Colonster, 20/43. B-4000 Liège (Belgique).
Univ. Liège. Faculté de Médecine vétérinaire. Service de Nutrition. Boulevard de Colonster, 20/43. B-4000 Liège (Belgique).
Résumé
Cette étude a pour but de quantifier les rejets azotés des vaches laitières au pâturage en rotation avec différents types de fertilisation (compost, lisier ou azote minéral), à des jours différents lors du séjour dans la parcelle (jours 3 et 5) pendant deux périodes différentes (juin et septembre). Des mesures de la production d'herbe et de la hauteur de l'herbe ont été réalisées. La composition chimique de l'herbe a été déterminée. Les animaux ont été pesés à l'entrée dans la parcelle et les productions laitières mesurées à chaque traite. Les ingestions d'azote ont été mesurées sur base des besoins des animaux. Lors du séjour dans les parcelles, des échantillons individuels de matières fécales et d'urine ont été prélevés afin de déterminer les rejets azotés. La teneur en urée dans le lait du tank et dans des échantillons individuels a été déterminée. L'ingestion d' azote a augmenté le jour 3 par rapport au jour 5 et au mois de septembre par rapport au mois de juin, mais n'a pas été influencée par la fertilisation. La quantité d'azote produit dans le lait n'a été influencée ni par la fertilisation, ni par le jour, ni par le mois. Les excrétions d'azote urinaire ont été significativement plus élevées dans le lot d'azote minéral que dans les deux autres lots. L'azote excrété par les matières fécales et les urines a été plus élevé au jour 3 qu'au jour 5. Au mois de juin, il y a eu moins d'azote excrété par les urines qu'au mois de septembre et plus d'azote excrété par les matières fécales. Des équations ont été calculées afin de prédire les excrétions d'azote par les urines de vaches laitières au pâturage à partir des teneurs en urée dans le lait d'un groupe de vaches et la composition en MAT de l'herbe.
Abstract
Influence of the fertilization type, the day in the paddock and the month on nitrogen rejections in dairy cows. The present study aims to quantify nitrogen (N) rejections in dairy cows on a rotational grazing system with different types of fertilization (compost, slurry and mineral N) at two different days in the paddock (days 3 and 5) and during two different periods (June and September). Grass yield and height were measured along with the chemical composition. The live weight was recorded when the cows moved into the paddock and milk yields was recorded at each milking. N intakes were measured on the basis of the energy requirement. When the cows were in the paddocks, individual samples of faeces and urine were collected to assess N rejections. The urea content in milk from the tank or from the individual cows was also measured. N intake was higher on day 3 than on day 5 and in September as compared with in June but was not influenced by the fertilization. The amount of N produced in milk was not influenced either by the fertilization, by the day on the paddock or by the month. The amount of urinary N was significantly higher in the N mineral group than in the two other groups. The N excretion in faeces and urines was higher on day 3 than on day 5. In June, the N excretion by the urine was lower than in September while the N excretion by the faeces was higher. Equations were calculated in order to estimate the N excretion by the urine in grazing dairy cows from the urea content in milk of a group of cows.
1. Introduction
1En Wallonie, les surfaces occupées par les prairies représentent 50 % de la surface agricole utile. D'un point de vue économique et environnemental, l'herbe pâturée est l'aliment le moins cher pour l'alimentation des bovins. Pour le grand public, le pâturage est perçu positivement et constitue un environnement naturel favorable au bien-être animal.
2Dans les prairies permanentes composées principalement de graminées et de trèfle blanc, les ingestions d'azote (N) peuvent excéder les besoins des animaux. Cette ingestion d'azote en excès peut entrainer des rejets azotés élevés (Groff et al., 2005) qui sont éliminés dans les urines. Dans les urines, l'urée constitue la plus grande partie de l'azote urinaire. Au niveau des pissats, l'azote non utilisé pour la croissance des plantes peut être perdu par volatilisation, lessivage ou dénitrification. Dans les matières fécales, l'azote est présent surtout sous forme organique et les pertes par volatilisation à partir des matières fécales sont par contre insignifiantes (Petersen et al., 1998). Les pertes d'azote issues des élevages bovins restent un problème important pour l'environnement. L'Union européenne désire diminuer ces pertes. Dans le cadre de la Directive Nitrates (91/676EC), un code de bonnes pratiques a été édicté dans chaque état membre. En Belgique, un Programme de Gestion Durable de l'Azote (PGDA) a été initié. La quantité d'azote provenant des déjections et des engrais de ferme est limitée à 230 kg sur les prairies et à 115 kg sur les terres de culture.
3Les agriculteurs doivent aussi respecter un taux de liaison au sol (Lambert et al., 2010) qui tient compte de la production d'azote dans leur élevage et de leur surface d'épandage. Les engrais de ferme sont donc pris en compte dans les plans de fumure. Ils peuvent conduire à une faible efficacité de l'utilisation et à des pertes d'azote en raison des émissions (Menzi et al., 1997 ; Stevens et al., 1997). Il existe peu d'études comparant l'efficacité des engrais de ferme, comme le compost et le lisier, et de l'azote minéral sur l'efficience de l'azote au pâturage.
4Meura et al. (2007), dans des prairies pâturées en Wallonie, ont rapporté que la teneur en matières azotées totales (MAT) de l'herbe augmentait au cours de la saison de pâturage, tandis que la teneur en sucres solubles diminuait. Cette quantité d'azote dépasse les capacités de prélèvement pour la production de l'herbe, surtout en fin de saison de pâturage où la croissance est réduite.
5Les vaches laitières pâturent habituellement dans un système en rotation où la qualité et la quantité d'herbe varient au cours du séjour dans la parcelle (Lopes et al., 2004). Ces modifications sont susceptibles d'influencer l'ingestion de matière sèche et les rejets azotés.
6Les rejets azotés peuvent être estimés à partir des teneurs en urée dans le lait (Kohn et al., 2002 ; De Campeneere et al., 2006) qui peuvent être déterminées par une méthode simple et peu couteuse. Il existe peu d'études avec des animaux au pâturage, malgré que les animaux y passent presque la moitié de l'année. Au pâturage, les paramètres expérimentaux sont difficiles à contrôler et les données sont plus variables que celles obtenues à l'étable (Schellberg et al., 2004).
7Cette étude a pour but de quantifier les rejets azotés chez des vaches laitières au pâturage en rotation avec différents types de fertilisation (compost, lisier ou azote minéral), à des jours différents lors du séjour dans la parcelle (jours 3 et 5) et pendant deux périodes différentes (juin et septembre).
2. Matériel et méthodes
8Les expériences ont eu lieu à la Station expérimentale de l'Université de Liège dans des prairies permanentes non drainées (50°33'N, 5°35'W). Le sol est constitué d'argile (< 0,002 mm : 17,3 %), de limon (0,002-0,05 mm : 71,6 %) et de sable (0,05-2 mm : 11,1 %). La présente étude a été réalisée pendant les années 5 et 6 d'une expérimentation testant les effets de trois types de fertilisation sur le bilan azoté de la prairie et les reliquats en nitrate dans le sol. Le protocole expérimental a été décrit par Dufrasne et al. (2007).
2.1. Protocole expérimental et animaux
9Les essais ont eu lieu à deux périodes différentes pendant la saison de pâturage en septembre et en juin l'année suivante. Ces mesures ont été prises en juin sur 35 vaches Holstein ayant vêlé depuis 164 ± 93 jours et en septembre sur 15 vaches Holstein ayant vêlé depuis 254 ± 72 jours. La semaine avant les essais, leur production laitière était 18,5 ± 3,7 kg par jour en juin et de 14,9 ± 4,7 kg par jour en septembre. Elles ont reçu 1 kg de pulpes séchées par jour lors de la traite.
2.2. Gestion du pâturage
10Dans le cadre du protocole appliqué pendant six ans, les vaches ont disposé de six parcelles pâturées en rotation. Les parcelles ont été pâturées quatre à cinq fois par saison. Les périodes de pâturage ont commencé au début du mois de mai et se sont terminées en octobre. Le chargement sur toute la saison de pâturage a été de 2,4 vaches par ha. Trois des parcelles ont été utilisées pour les essais présentés dans la présente publication. La durée de séjour lors des essais a été de sept jours. Les parcelles étaient pâturées pour la deuxième fois et pour la quatrième fois dans le cycle de rotation lors des essais de juin et de septembre respectivement. Les chargements ont été de 12,8 vaches par ha et 11,2 vaches par ha en juin et en septembre respectivement. La première parcelle pâturée était amendée avec de l'engrais azoté minéral (Nmin), la deuxième avec du lisier de porc (L) et la troisième avec du compost (C). La durée de repousse entre deux cycles de rotation a été de 33 jours en juin et de 28 jours en septembre. La surface offerte par vache a été de 111 m2 en juin et 127 m2 en septembre.
11Dans la parcelle Nmin, la première fertilisation au printemps a apporté du P2O5 et K2O. Avant l'essai de juin, les apports totaux de N, P2O5 et K2O par la fertilisation ont été de 56, 18 et 36 kg.ha-1 respectivement dans la parcelle Nmin ; 69, 31, 54 kg.ha-1 dans la parcelle L ; 78, 52 et 78 kg.ha-1 dans la parcelle C.
12Avant l'essai réalisé au mois de septembre, les apports totaux de N, de P2O5 et de K2O ont été de 78, 36 et 36 kg.ha-1 dans la parcelle Nmin ; 105, 47, 60 kg.ha-1 dans la parcelle L et 105, 52 et 78 kg.ha-1 dans la parcelle C.
13La prairie était composée de ± 85 % de graminées, représentées principalement par le ray-grass anglais, de 10 % de trèfle blanc et de 5 % d'autres plantes.
2.3. Mesures phytotechniques
14Les productions d'herbe ont été déterminées avant l'entrée dans la parcelle sur trois bandes de dix mètres de long à l'aide d'une tondeuse. Les hauteurs d'herbe ont été mesurées à l'aide d'un herbomètre constitué d'une plaque d'aluminium de 30 cm de côté excerçant une pression de 2 kg.m-2 coulissant sur un pied gradué. Les mesures ont été réalisées avant l'entrée des animaux dans la parcelle et à la sortie.
15Des échantillons d'herbe représentant 10 à 15 poignées par parcelle ont été prélevés aux jours 2 et 4, sèchés à l'étuve et broyés. Les cendres, la matière azotée totale (MAT), les fractions NDF (fibres au détergent neutre) et ADF (fibres au détergent acides), les sucres solubles et les UFL ont été déterminés par spectrométrie dans le proche infrarouge.
2.4. Mesures sur les animaux
16Les vaches ont été traites deux fois par jour à 6h00 et à 15h30. Les productions de lait ont été mesurées à chaque traite. Dans l'essai du mois de septembre, un échantillon de lait dans le tank a été prélevé à chaque traite pour déterminer les teneurs en azote et en urée. Dans l'essai du mois de juin, des échantillons individuels de lait ont été prélevés le matin et l'après-midi et un échantillon représentant la traite de la journée a été constitué. Un échantillon d'urine obtenu par stimulation manuelle de la vulve et un échantillon de matières fécales collecté dans le rectum ont été prélevés sur chaque vache entre 9h00 et 10h00 les 3e et 5e jours lors du séjour sur les parcelles. Les échantillons ont été divisés en deux. Un des échantillons a été utilisé pour la détermination de la créatinine réalisée le jour même. L'autre échantillon a été congelé à -20 °C. Les échantillons de matières fécales ont été sèchés à l'étuve et broyés. Les vaches ont été pesées à 10h00 le jour des prélèvements à l'aide d'une balance électronique.
2.5. Mesure du bilan azoté
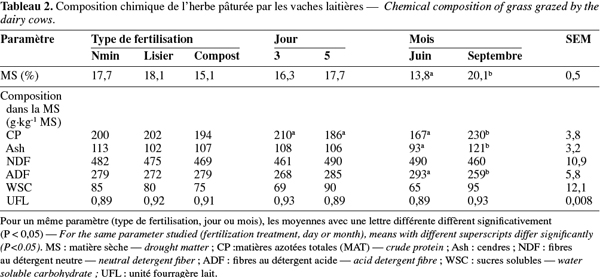
17Les besoins en énergie exprimés en UFL pour l'entretien, le pâturage, la production laitière et la gestation ont été calculés selon Jarrige (1980) :
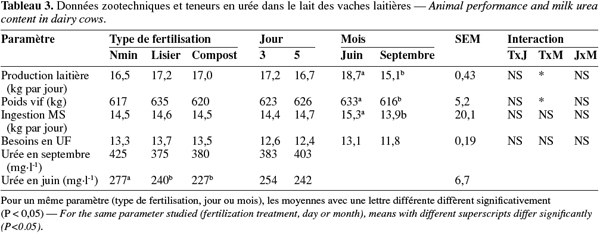
18UFL entretien = 1,4 + 0,006 p où p est le poids en kg
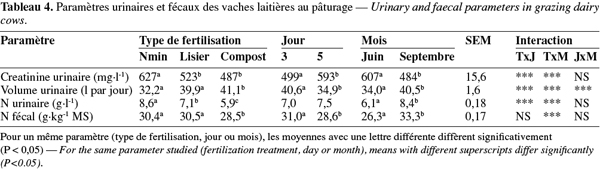
19UFL pâturage = 0,10 x UFL entretien
20UFL production laitière = 0,4 x l lait à 3,5 % matière grasse
21UFL gestation = + 1 pour le 7e mois, + 1,5 pour le 8e mois et + 2 pour le 9e mois
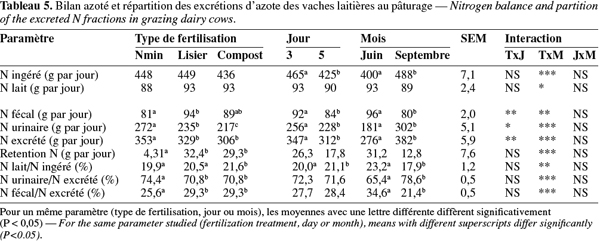
22Total besoins énergétiques en UFL = UFL entretien + UFL pâturage + UFL production laitière + UFL gestation
23L'énergie apportée par l'herbe a été calculée par différence entre les besoins énergétiques totaux et l'énergie apportée par le complément. L'ingestion d'herbe exprimée en kg de matière sèche (MS) a été obtenue en divisant l'énergie apportée par l'herbe ingérée divisée par la densité énergétique de l'herbe. L'ingestion d'azote est la somme de l'azote apporté par les compléments et par l'herbe.
24La créatinine urinaire, secrétée proportionnellement à la masse musculaire à raison de 100 mg.kg-1, a été utilisée pour estimer le volume urinaire quotidien (Marini et al., 2005). Selon Clinquart et al. (1994) et Minet et al. (1996), le rendement à l'abattage des carcasses de vaches laitières est de 50,7 % et la proportion de muscles dans la carcasse, de 54,2 %. La quantité d'azote excrété par les urines a été calculée à partir du volume urinaire et de la concentration en azote dans les urines. La détermination des cendres insolubles dans l'herbe, dans le complément et les matières fécales, a été utilisée pour déterminer l'excrétion d'azote dans les matières fécales. La teneur en azote du lait a été obtenue en divisant la quantité de matière azotée de lait par 6,38. La rétention d'azote a été calculée par soustraction de la quantité d'azote excrété par les matières fécales, les urines et le lait de l'ingestion de l'azote total.
2.6. Analyses
25La créatinine a été déterminée dans l'urine fraiche par la méthode colorimétrique de Jaffe. Le contenu en azote a été déterminé dans les urines congelées et dans les matières fécales par la méthode de Kjeldahl. Les cendres, la MAT, les fractions NDF, ADF et les sucres solubles de l'herbe ont été mesurés par spectrométrie dans le proche infrarouge. Les cendres insolubles ont été mesurées dans les matières fécales, l'herbe et les pulpes séchées par la méthode de Van Keulen et Young (1977). Les taux de matière grasse, de matière protéique et les teneurs en urée dans le lait ont été analysés par spectrométrie dans le moyen infrarouge (Foss Milkoscan FT6000 ; Foss, Hillerod, Denmark).
2.7. Analyses statistiques
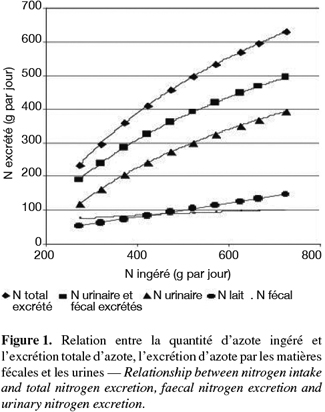
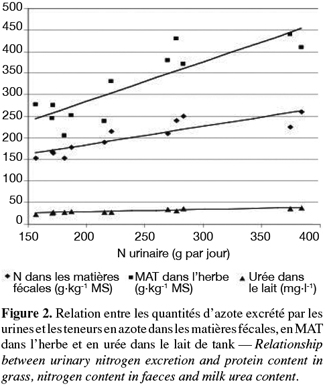
26Les productions d'herbe et les hauteurs d'herbe ont été analysées par la procédure general linear model du logiciel Minitab (1989). Le modèle statistique suivant a été utilisé : Yij = µ + αi + βj où µ = moyenne, αi = effet fixe du traitement fertilisation et βj = effet fixe du mois.
27Les autres données phytotechniques et les données concernant les animaux ont été analysées à l'aide d'un modèle mixte (proc mixed, SAS, 1999) incluant une autocorrélation entre des mesures successives sur des animaux identiques à l'intérieur d'un traitement. Les effets des traitements, du jour de récolte, du mois et les interactions entre ces paramètres ont été inclus dans le modèle.
3. Résultats
3.1 Conditions climatiques
28Les précipitations en juin ont été similaires à la moyenne des dix dernières années, soit 72 l. Les précipitations en septembre ont été inférieures de 12 l par rapport à la moyenne des dix dernières années qui était de 64 l.
29Le mois de juin avec une température moyenne de 15,6 °C a été plus froid que la moyenne des dix dernières années, qui était de 16,6 °C. La température moyenne en septembre a été de 18,8 °C, alors que la moyenne des dix dernières années a été de 15,4 °C.
3.2. Mesures phytotechniques
30Les hauteurs à l'entrée de la parcelle ont eu tendance à être plus élevées dans la parcelle Nmin (Tableau 1 : 15,1 cm) que dans les parcelles L et C (14,6 et 13,1 cm respectivement). Les hauteurs à la sortie ont été de 6,2 cm en moyenne et n'ont pas été différentes en fonction des traitements. La production d'herbe a été plus élevée dans la parcelle Nmin (2 740 vs en moyenne 2 177 kg MS.ha-1 ; NS). La hauteur de l'herbe à l'entrée a été plus élevée en juin qu'en septembre (16,1 vs 12,5 cm ; P < 0,05), mais il n'y a pas eu de différence pour la hauteur de sortie. La production d'herbe a été plus élevée en juin qu'en septembre (3 188 vs 1 545 kg MS.ha-1 ; P < 0,05).
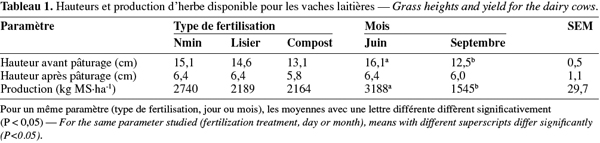
31Le type de fertilisation n'a pas eu d'effet sur la composition en MAT, cendres, NDF, ADF, en sucres solubles et en UFL (Tableau 2). Le jour de prélèvement a eu un effet significatif uniquement sur la teneur en MAT qui a été plus haute le jour 3 par rapport au jour 5 (P < 0,05). Le mois a eu une influence significative sur les teneurs en MAT et en cendres qui ont été plus élevées en septembre (230 vs 167 g CP.kg-1 MS, P < 0,01 ; 121 vs 93 g.kg-1 MS, P < 0,05 respectivement). Par contre, la composition en ADF a été significativement plus élevée en juin qu'en septembre (293 vs 259 g.kg-1 MS, P < 0,05).
3.3. Performances des animaux et teneurs en urée dans le lait
32Les poids vif et la production laitière n'ont pas été influencés par le traitement, ni par le jour de prélèvement, mais bien par le mois (Tableau 3). Les vaches étaient plus lourdes et ont produit plus de lait en juin qu'en septembre (633 vs 616 kg et 18,7 vs 15,1 kg par jour, P < 0,05 et P < 0,001). L'ingestion de matière sèche et les besoins énergétiques exprimés en UFL par vache n'ont pas été affectés par le traitement, ni le jour de récolte, mais ont été plus élevés en juin qu'en septembre (P < 0,001). Les teneurs en urée déterminées individuellement dans le lait en juin ont été significativement plus élevées dans le lot Nmin que dans le lot L et C (277 vs 240 et 227 mg.l-1, P < 0,01). Le jour de récolte n'a pas eu d'effet.
3.4. Bilan azoté
33La teneur en créatinine et le volume urinaire ont été influencés par le type de fertilisation, le jour de récolte et le mois (Tableau 4, P < 0,001). Le volume urinaire a été plus faible dans le lot Nmin qu'avec les autres types de fertilisation, au jour 5 qu'au jour 3 et en juin qu'en septembre (32,2 vs 40,6 ; 34,9 vs 40,6 ; 34,0 vs 40,5 l par jour respectivement, P < 0,001). Les teneurs en azote dans les urines et les matières fécales ont aussi été influencées par le type de fertilisation (P < 0,001). La teneur en azote dans les urines a été plus élevée dans le lot Nmin et la plus faible dans le lot C (8,6 vs 5,9 g N.l-1, P < 0,01). La teneur en azote dans les matières fécales a été plus élevée dans le lot L et dans le lot Nmin que dans le lot C (30,5 et 30,4 vs 28,5 g N.kg-1 MS respectivement, P < 0,001). Le jour de récolte n'a pas influencé la teneur en azote urinaire. La teneur en azote fécal a été plus élevée au jour 3 qu'au jour 5 (31,0 vs 28,6 g N.kg-1 MS, P < 0,01). Les teneurs en azote urinaire et fécal ont été plus élevées en septembre qu'en juin (8,4 vs 6,1 g N.l-1 ; 33,3 vs 26,3 g N.kg-1 MS respectivement, P < 0,001). Il y a eu des interactions significatives entre le traitement et le mois (P < 0,001).
34L'ingestion d'azote a été similaire avec les trois fertilisations (444 g par jour, en moyenne) mais a été significativement plus élevée au jour de récolte 3 qu'au jour de récolte 5 (Tableau 5 : 465 vs 425 g par jour, P < 0,001) et en septembre qu'en juin (488 vs 400 g par jour, P < 0,01). La quantité d'azote produit dans le lait n'a pas été différente selon le type de fertilisation, le jour de récolte ou le mois (91 g par jour, en moyenne). L'excrétion d'azote par les matières fécales a été significativement influencée par la fertilisation, le mois et le jour de récolte (P < 0,001). Elle a été plus élevée dans le lot L qu'avec les lots C et Nmin (94 vs 89 vs 81 g N par jour). Les valeurs pour le lot C n'ont pas été significativement différentes des deux autres lots. L'excrétion d'azote par les matières fécales a été plus élevée au jour 3 qu'au jour 5 (92 vs 84 g N par jour, P < 0,01) et en juin qu'en septembre (96 vs 80 g N par jour, P < 0,001). Il y a eu des interactions significatives entre le type de fertilisation et le jour de récolte d'une part (P < 0,01) et le mois d'autre part (P < 0,01).
35L'excrétion d'azote par les urines a été plus importante dans le lot Nmin que dans les lots L et C (272 vs 235 et 217 g par jour, P < 0,001). Les vaches ont excrété plus d'azote dans les urines au jour de récolte 3 qu'au jour 5 (256 vs 228 g par jour, P < 0,001) et en septembre qu'en juin (302 vs 181 g par jour, P < 0,001). Il y a eu des interactions significatives entre le type de fertilisation et le jour d'une part (P < 0,05) et le mois d'autre part (P < 0,001). Les quantités d'azote excrété par les urines et les matières fécales ont été significativement plus élevées dans le lot Nmin que dans les lots L et C (353 vs 329 et 306 g par jour, P < 0,01) au jour de récolte 3 qu'au jour de récolte 5 (347 vs 312 g par jour, P < 0,01) et en septembre qu'en juin (382 vs 276, P < 0,001). Il y a eu des interactions significatives entre le type de fertilisation et le jour d'une part (P < 0,01) et le mois d'autre part (P < 0,001). La rétention d'azote a été significativement plus faible dans le lot Nmin qu'avec les autres fertilisations (4,31 vs 32,4 et 29,3 g par jour , P < 0,05) mais il n'y a pas eu de différence significative pour le jour de récolte et le mois.
36La proportion d'azote produit dans le lait par rapport à l'azote total ingéré a été plus élevée dans le lot C, au jour 5 et en juin. La proportion d'azote excrété par les urines par rapport à l'azote total excrété a été plus élevée dans le lot Nmin que dans les autres lots (74,4 vs 70,8 %, P < 0,001) et en septembre (78,6 vs 65,4 %, P < 0,001), mais n'a pas été influencée par le jour de récolte. Des interactions significatives ont été calculées entre le type de fertilisation et le mois pour toutes les variables étudiées.
37Des relations curvilinéaires et linéaires positives ont été mises en évidence entre les quantités d'azote ingéré d'une part et les quantités d'azote excrété par les urines, les matières fécales, le lait et l'excrétion totale d'azote composée de l'azote des matières fécales, de l'azote urinaire et de l'azote du lait (Figure 1) :
38N urinaire (g par jour) = 281,5 ln N ingéré (g par jour) - 1459,6 ; r2 = 0,34, P < 0,0001
39N fécal (g par jour) = 0,062 x N ingéré (g par jour) + 58,8 ; r2 = 0,06, P < 0,0001
40N lait (g par jour) = 0,21 x N ingéré (g par jour) - 3,8 ; r2 = 0,56, P < 0,0001
41N urinaire (g par jour) + N fécal (g par jour) = 310,4 ln N ingéré (g par jour) - 1549,0 ; r2 = 0,41, P < 0,0001
42Excrétion totale d'N = 404,4 ln N ingéré (g par jour) - 2032,1 ; r2 = 0,57, P < 0,0001
43L'excrétion d'azote urinaire pour un lot de vaches peut être prédite à partir de la composition en MAT de l'herbe, de la teneur moyenne en urée dans le lait ou de la composition moyenne en azote fécal (Figure 2). Des relations linéaires ont été calculées :
44N urinaire (g par jour) = -113 + 1,77 MAT de l'herbe (g.kg-1 MS), r2 = 0,74, p < 0,001 (équation 1)
45N urinaire (g par jour) = -20,7 + 0,83 urée dans le lait (mg.l-1), r2 = 0,73, p < 0,001 (équation 2)
46N urinaire (g par jour) = -241 + 15,3 teneur en N fécal (g.kg-1 MS), r2 = 0,82, p < 0,001(équation 3)
47N urinaire (g par jour) = -183 + 9,51 teneur en N fécal (g.kg-1 MS) + 0,19 MAT de l'herbe (g.kd-1 MS) + 0,32 urée dans le lait (mg.l-1), r2 = 0,86, P < 0,001
4. Discussion
4.1. Données phytotechniques
48La hauteur de l'herbe à la sortie des parcelles n'a pas été considérée comme une valeur limitante pour la production de lait (Hoden et al., 1991). La hauteur de l'herbe à l'entrée des parcelles et la production d'herbe ont eu tendance à être plus élevées dans la parcelle Nmin. Il faut cependant noter que sur une durée de six ans, Dufrasne et al. (2007) ont rapporté une production plus élevée dans la parcelle L que dans la parcelle C, la parcelle Nmin ayant présenté des valeurs intermédiaires. La composition en MAT, cendres, NDF, ADF et UFL n'a pas été différente entre les traitements de fertilisation comme rapporté par Dufrasne et al. (2007) sur l'ensemble des six ans. La hauteur de l'herbe et la production d'herbe plus élevées en juin qu'en septembre sont dues à un effet saisonnier de la croissance de l'herbe. L'augmentation de la teneur en MAT en septembre peut être la conséquence d'un temps de repousse plus court (28 vs 33 jours) comme l'ont rapporté Owens et al. (2008). Meura et al. (2007) ont signalé aussi une augmentation de la teneur en MAT de l'herbe au cours de la saison de pâturage. La teneur plus faible en MAT au jour 5 par rapport au jour 3 peut être attribuée à un pâturage sélectif, les feuilles plus riches en MAT que les tiges étant d'abord ingérées.
4.2. Performances zootechniques et besoins en énergie
49Comme l'ont rapporté Dufrasne et al. (2007) dans l'essai sur six années, la production laitière au cours de la saison de pâturage n'a pas été influencée par le type de fertilisation. La diminution de la production de lait au jour 5 par rapport au jour 3 peut être expliquée par la conduite du pâturage. Dans un pâturage en rotation, quand les vaches entrent dans une nouvelle parcelle, la disponibilité en herbe est élevée et la production laitière augmente pour atteindre un maximum. A la fin du séjour dans la parcelle, la production laitière diminue car la disponibilité en herbe devient faible. L'effet significatif du mois sur la production laitière peut être expliqué par le stade de lactation et la disponibilité en herbe : en septembre, les vaches étaient en fin de lactation et la production d'herbe était plus faible qu'en juin. La production laitière et le poids vif plus élevé en juin expliquent les besoins accrus en énergie.
4.3. Bilan azoté
50La teneur en urée qui reflète les teneurs en ammoniaque dans le rumen peut être considérée comme un indicateur des rejets azotés (Jonker et al., 1998). Les valeurs moyennes en juin de 248 mg.l-1 et en septembre de 393 mg.l-1 ont été plus faibles que celles rapportées par Dieguez et al. (2001) qui étaient de 401 mg.l-1 pour l'ensemble de la saison. De Brabander et al. (1999) considèrent que la teneur en urée dans le lait est satisfaisante lorsqu'elle est comprise entre 175 et 300 mg.l-1. Les valeurs de septembre sont trop élevées par rapport à cette norme, mais au pâturage, Paccard et al. (1999) considèrent que les valeurs repères peuvent dépasser 400 mg.l-1. Certains auteurs considèrent que des teneurs élevées en urée et une alimentation trop riche en protéines peuvent occasionner des problèmes au niveau de la sphère génitale (Ferguson et al., 1993 ; Rajala-Schultz et al., 2001). Par contre, Guo et al. (2004) n'ont pas signalé d'effet des teneurs en urée dans le lait sur la fécondité. Dufrasne et al. (2008), dans une étude sur les effets des facteurs environnementaux sur les teneurs en urée dans le lait dans les exploitations laitières wallonnes pendant six ans, ont rapporté une moyenne de 282 mg.l-1 d'urée en juin et de 343 mg.l-1 en septembre. Ils ont expliqué les différences obtenues entre les mois par le changement de composition de l'herbe : une augmentation de la teneur en MAT et une diminution de la teneur en sucres solubles étant observées pendant la saison de pâturage. De tels changements peuvent conduire à un déséquilibre alimentaire en fin de saison de pâturage provoquant des teneurs en urée élevées. Ces modifications au niveau de la composition de l'herbe ont également été observées dans le présent essai.
51Les teneurs en urée du lait de tank relevées en septembre et les teneurs en urée individuelles mesurées en juin ont montré toutes deux des valeurs plus élevées dans le lot Nmin. Cette observation ne peut pas être expliquée par une différence de MAT au niveau de l'herbe. Dans les prairies pâturées par des vaches laitières, Dieguez et al. (2001) ont signalé des teneurs en urée dans le lait plus élevées dans des prairies fertilisées avec Nmin que dans des pâtures sans fertilisation, alors que la composition en MAT était identique. Ils ont attribué cette différence au fait que la teneur en nitrate de l'herbe était plus élevée dans les parcelles Nmin. La teneur en nitrate n'a pas été mesurée dans cet essai mais pourrait être affectée par le type de fertilisation. Les teneurs en urée dans le lait plus élevées en septembre qu'en juin peuvent s'expliquer par la teneur en MAT de l'herbe supérieure en septembre.
52Les quantités d'azote ingéré ont été supérieures à celles citées par Steinshamm et al. (2006) avec des vaches laitières au pâturage, mais la teneur en MAT de l'herbe a été plus élevée dans cet essai. Mulligan et al. (2004) ont obtenu des ingestions similaires quand les vaches recevaient 1 kg de concentré riche en protéines ou 6 kg de concentré faible en protéines. Les quantités d'azote ingéré supérieures au jour 3 par rapport au jour 5 et en septembre par rapport à juin sont le résultat de l'ingestion d'une herbe plus riche en MAT.
53L'azote produit dans le lait a été similaire aux valeurs observées par Steinshamn et al. (2006) et reflète une production normale pour des vaches au pâturage à ce stade de lactation recevant peu de concentré. Il y a eu une relation étroite entre l'azote produit dans le lait et les quantités d'azote ingéré. La proportion d'azote produit dans le lait par rapport à l'azote ingéré (20,6 %) a été plus basse que celle obtenue par Mulligan et al. (2004) et par Steinshamn et al. (2006) mais correspond aux valeurs renseignées par Burke et al. (2008). La faible efficacité de l'utilisation de l'azote dans le lot Nmin corrobore les teneurs en urée plus élevées observées dans ce lot. L'efficacité plus élevée au jour 5 est liée à de plus faibles ingestions d'azote pour une production de lait similaire. La meilleure efficience en juin par rapport à septembre peut être expliquée par de plus faibles ingestions d'azote et une production laitière supérieure.
54La teneur en azote dans les matières fécales, similaire à celles citées par Leonardi et al. (2003), a augmenté au jour 3 et en septembre quand l'ingestion d'azote était plus élevée. Ces résultats sont en accord avec ceux de Leonardi et al. (2003) qui ont comparé deux niveaux de MAT dans des rations distribuées à l'étable. Schellberg et al. (2007) ont rapporté que la teneur en azote des matières fécales et de l'herbe ont évolué dans le même sens et ont conclu que la teneur en azote dans les matières fécales peut être utilisée comme indicateur de la composition en azote de l'herbe de prairies qui présentent des variations de composition ou de production de MS. Dans la présente étude, la relation entre la teneur en azote des matières fécales et de l'herbe a été de 72 %. La teneur en azote dans les matières fécales a également montré une meilleure relation avec les quantités d'azote ingéré (r2 = 0,22, non montré) que les quantités d'azote excrété par les matières fécales (r2 = 0,06). L'excrétion moyenne d'azote par les matières fécales a été de 88 g par jour ; cette valeur est plus basse que celle calculée par Mulligan et al. (2004), mais est en concordance avec les résultats de Schellberg et al. (2004) obtenus avec des vaches allaitantes et ceux de Steinshamn et al. (2006) qui ont mesuré des excrétions variant de 76 jusqu'à 122 g. Peyraud et al. (1995) ont rapporté une excrétion d'azote par les matières fécales de 7,5 g.kg-1 MS ingérée ; avec une moyenne de 14,4 kg MS ingérée, l'excrétion d'azote par les matières fécales aurait dû être de 108 g par jour. Il est possible que l'excrétion d'azote par les matières fécales ait été sous-estimée. Cela pourrait expliquer les valeurs élevées de rétention d'azote qui ont été en moyenne de 5 % par rapport aux quantités d'azote ingéré (22 g en moyenne / 444 g en moyenne, tableau 5). La rétention d'azote n'est pas souvent calculée dans les essais de pâturage, les valeurs d'excrétion de l'azote total étant habituellement obtenues par différence des quantités d'azote ingéré et de l'azote produit dans le lait. La rétention d'azote peut être considérée comme normale : Gibb et al. (1992) citent une rétention d'azote de 26 g.kg-1 de gain de poids et Leonardi et al. (2003) ont observé une rétention d'azote de 68 g par jour avec une ingestion d'azote de 635 g par jour. Dans un pâturage en rotation, il n'est pas possible de distinguer les changements de poids vif dus au remplissage des estomacs des gains de poids des muscles. De plus, les besoins en azote pour la gestation des vaches doivent être pris en compte : ils sont estimés entre 4,0 et 9,8 g par jour pour des vaches en seconde partie de lactation (Jarrige, 1980). La faible relation entre l'excrétion d'azote par les matières fécales et les quantités d'azote ingéré (r2 = 0,06) est en concordance avec les résultats de Schellberg et al. (2007) avec des vaches allaitantes au pâturage et de Leonardi et al. (2003) avec des vaches laitières à l'étable. Par contre, Mulligan et al. (2004) ont montré que l'excrétion d'azote par les matières fécales augmentait en même temps que l'ingestion d'azote.
55Les volumes urinaires ont été plus élevés que ceux mesurés par Leonardi et al. (2003) sur des vaches laitières à l'étable recevant une ration dont la teneur en MS était de 55 %. Dans la présente étude, la ration était composée principalement d'herbe avec une teneur en MS de 13,8 % en juin et de 20,1 % en septembre ; la faible teneur en MS peut expliquer les volumes urinaires plus élevés. De plus, Leonardi et al. (2003) ont constaté une augmentation du volume urinaire lorsque la teneur en MAT de la ration était augmentée. Cette observation a également été réalisée dans la présente étude avec des volumes urinaires plus élevés en septembre, malgré l'ingestion d'une herbe plus riche en MS. L'excrétion d'azote par les urines a été en moyenne de 242 g par jour et était la principale source d'excrétion d'azote. Ces résultats sont en concordance avec ceux rapportés dans la littérature pour des vaches au pâturage. La relation avec l'excrétion totale d'azote a été bonne (r2 = 0,88). La quantité d'azote excrété par les urines et la proportion d'azote excrété par les urines par rapport à l'azote total excrété, plus élevées dans le lot Nmin, confirment les teneurs en urée plus élevées observées dans ce lot. Par contre dans le lot C, les quantités plus faibles d'azote excrété par les urines sont en concordance avec les teneurs en azote des matières fécales et des urines ; les teneurs en urée dans le lait étaient également plus basses dans ce lot. Une analyse plus fine des fractions azotées non réalisée dans cette étude pourrait probablement expliquer ces résultats. L'excrétion d'azote par les urines plus élevée au jour 3 qu'au jour 5 peut s'expliquer par les ingestions d'azote plus élevées. L'azote excrété par les urines a chuté de 22 g N.kg-1 MS ingéré en septembre à 12 g N.kg-1 MS ingéré en juin ; ces résultats sont en accord avec ceux de Peyraud et al. (1998) obtenus lorsque la teneur en MAT de l'herbe diminue suite à une fertilisation azote moindre ou à une durée de repousse plus longue. Les quantités d'azote excrété par les urines et la proportion d'azote excrété dans les urines par rapport à l'azote total excrété, plus élevées en septembre qu'en juin, ainsi que la faible efficience de l'utilisation de l'azote dans le lait en septembre indiquent que les ingestions d'azote ont été trop élevées par rapport aux besoins des vaches qui étaient en fin de lactation. Selon le NRC (2001), pour des vaches laitières recevant une ration à base de fourrages, les besoins en MAT sont compris entre 160 et 180 g MAT.kg-1 MS. Les valeurs en MAT dans l'herbe plus basses en juin (167 g.kg-1 MS) qu'en septembre (230 g.kg-1 MS) correspondent mieux aux besoins des vaches. Le concentré énergétique (pulpes sèchées) était apporté en trop faible quantité (1 kg par jour) pour compenser les excès de MAT ingérés au mois de septembre. Il n'est pourtant par rare, dans les exploitations commerciales, qu'un complément riche en MAT soit apporté à de l'herbe pâturée en fin de saison de pâturage afin de soutenir la production laitière. Dans ces conditions, l'ingestion d'azote est encore augmentée, ce qui peut provoquer une augmentation de l'azote excrété. Il est donc indispensable que la complémentation soit bien adaptée en fonction de la composition de l'herbe.
56Il est utile de disposer d'équations permettant d'estimer la quantité d'azote excrété par les urines à partir de données facilement disponibles comme les teneurs en urée dans le tank et la teneur en MAT de l'herbe afin de pouvoir contrôler les pertes en azote. Au pâturage, on dispose de peu de données. Dans cette étude, des relations étroites entre les excrétions d'azote par les urines et les teneurs en urée dans le lait d'un groupe de vaches ou avec la teneur en MAT ont été mises en évidence (équations 1 et 2). La prédiction est améliorée quand les paramètres sont utilisés ensemble. Afin d'estimer plus précisément les excrétions d'azote par les urines au pâturage, la teneur en azote dans les matières fécales qui a montré une meilleure corrélation avec l'excrétion d'azote par les urines (équation 3) peut être proposée, les matières fécales étant plus faciles à collecter que les urines.
5. Conclusion
57Cette étude a permis d'obtenir des données sur l'excrétion d'azote chez des vaches laitières dans un pâturage en rotation avec différents types de fertilisation, à différents moments lors du séjour dans la parcelle et à différentes périodes de la saison de pâturage. Les types de fertilisation ont influencé les excrétions d'azote composées principalement d'azote urinaire. En fin de saison de pâturage, des excrétions élevées liées à une teneur en MAT de l'herbe augmentée ont été mises en évidence. Il convient que la nature de la complémentation de l'herbe à ce moment soit définie avec pertinence afin de ne pas aggraver les pertes en azote. A partir de paramètres facilement disponibles comme les teneurs en urée dans le lait ou la composition en MAT de l'herbe, il est possible de prédire les excrétions d'azote chez des vaches au pâturage. Des recherches supplémentaires sont nécessaires afin de préciser l'influence de la fertilisation et de la composition de l'herbe au cours de la saison de pâturage sur l'excrétion d'azote.
Bibliographie
Burke F. et al., 2008. Effect of pasture allowance and supplementation with maïze silage and concentrate differing in crude protein concentration on milk production and nitrogen excretion by dairy cows. Livest. Sci., 114, 325-335.
Clinquart A. et al., 1994. Meat quality in relation to breed (Belgian Blue vs Hostein) and conformation (double muscled vs dual purpose type). Sci. Aliments, 14, 403-409.
De Brabander D. et al., 1999. Invloedsfactoren op het melkureumgehalte. Informatienamiddag: een doelbewuste melkveevoeding – goed voor de boer, het dier en het milieu. Gent, Belgie: Ministerie van Middenstand en Landbouw, Department dierenvoeding en veehouderij.
De Campeneere S., de Brabander D.L. & Vanacker J.M., 2006. Milk urea concentration as affected by the roughage type offered to dairy cattle. Livest. Sci., 103, 30-39.
Dieguez F., Hornick J.-L., Istasse L. & Dufrasne I., 2001. Désintensification raisonnée de l'exploitation au pâturage par la vache allaitante Blanc Bleu Belge. Fourrages, 165, 61-71.
Dufrasne I. et al., 2007. Nitrogen balance and nitrate residues in pastures grazed by dairy cows and fertilised with mineral fertiliser, pig slurry or cattle compost. In: Proceedings of the European Grassland Federation, 3-5 September, Ghent, Belgium, 343-346.
Dufrasne I., Cabaraux J.-F., Istasse L. & Hornick J.-L., 2008. Milk urea content: effects of environmental parameters and relationships with other milk traits. In: Proceedings of 22nd General meeting of European Grassland Federation, 9-12 June, Sweden, 577-579.
Ferguson J.D., Galligan D.T., Blanchard T. & Reeves M., 1993. Serum urea nitrogen and conception rate: the usefulness of test information. J. Dairy Sci., 76, 3742-3746.
Gibb M.J., Ivings W.E., Dhanoa M.S. & Sutton J.D., 1992. Changes in body components of autumn-calving Hostein-Friesian cows over the first 29 weeks of lactation. Anim. Prod., 55, 339-360.
Groff E.B. & Wu Z., 2005. Milk production and nitrogen excretion of dairy cows fed different amounts of protein and varying proportions of alfalfa and corn silage. J. Dairy Sci., 88, 3619-3632.
Guo K., Russek-Cohen E., Varner M.A. & Kohn R.A., 2004. Effects of milk urea nitrogen and other factors on probability of conception of dairy cows. J. Dairy Sci., 87, 1878-1885.
Hoden A. et al., 1991. Simplified rotational grazing management of dairy cows: effect of rates of stocking and concentrate. J. Agric. Sci., 116, 417-428.
Jarrige R., 1980. Alimentation des ruminants. Paris : INRA.
Jonker J.S., Kohn R.A. & Erdman R.A., 1998. Using milk urea nitrogen to predict nitrogen excretion and utilization efficiency in lactating dairy cows. J. Dairy Sci., 81, 2681-2692.
Kohn R.A., Kalscheur K.F. & Russek-Cohen E., 2002. Evaluation of models to estimate urinary nitrogen and expected milk urea nitrogen. J. Dairy Sci., 85, 227-233.
Lambert R. et al., 2010. Vers une révision de la norme de production d’azote de la vache laitière : justification et conséquences sur le taux de liaison au sol des exploitations laitières. Biotechnol. Agron. Soc. Environ., 10(S1), 67-71.
Leonardi C., Stevenson M. & Armentano L.E., 2003. Effect of two levels of crude protein and methionine supplementation on performance on dairy cows. J. Dairy Sci., 86, 4033-4042.
Lopes F.C.F. et al., 2004. Effect of supplementation and resting period on forage quality and voluntary intake on elephantgrass grazed by lactating Holstein x Zebu cows. Arq. Bras. Med. Vet. Zootec., 56, 355-362.
Marini J.C. & van Amburgh M.E., 2005. Partition of nitrogen excretion in urine and feces of Holstein replacement heifers. J. Dairy Sci., 88, 1778-1784.
Menzi H. et al., 1997. Ammonia emissions following the application of solid manure to grassland. In: Jarvis S.C. & Pain B.P., eds. Gaseous nitrogen emissions from grasslands. Wallingford, UK: CAB International, 265-274.
Meura S. et al., 2007. Milk urea content as influenced by geographical area and season in Wallonia. In: Proceedings of the European Grassland Federation, 3-5 September, Ghent, Belgium, 375-377.
Minet V. et al., 1996. Stratégies d'engraissement du taurillon Blanc Bleu Belge culard : performances, qualité des carcasses et de la viande, approche métabolique et bilan économique. Liège, Belgique : Presses de la Faculté de Médecine vétérinaire de Liège.
Mulligan F.J. et al., 2004. Supplementary concentrate type affects nitrogen excretion of grazing dairy cows. J. Dairy Sci., 87, 3451-3460.
NRC, 2001. Nutrient requirements of dairy cattle. 7th ed. Washington: National Academy Press.
Owens D., McGee & Boland T., 2008. Effect of regrowth interval on intake, rumen, digestion and nutrient flow to the omasum in beef cattle. Anim. Feed Sci. Technol., 146, 21-41.
Paccard P. et al., 1999. Le taux d'urée du lait de troupeau : un repère pour l'alimentation. Aubière, France : Institut de l'Elevage.
Petersen S.O., Sommer S.G., Aes O. & Soegaard K., 1998. Ammonia loss from urine and dung of grazing cattle: effect of N intake. Atmos. Environ., 32, 295-300.
Peyraud J.L., Vérité R. & Delaby L., 1995. Rejets azotés chez la vache laitière : effets du type d'alimentation et du niveau de production des animaux. Fourrages, 142, 131-144.
Peyraud J.L. & Astigarraga L., 1998. Review of the effect of nitrogen fertilization on the chemical composition, intake, digestion and nutritive value of fresh herbage: consequences on animal nutrition and N balance. Anim. Feed Sci. Technol., 72, 235-259.
Rajala-Schultz P.J., Saville W.J., Frazer G.S. & Wittum T.E., 2001. Association between milk urea and fertility in Ohio dairy cows. J. Dairy Sci., 84, 482-489.
Schellberg J. & Gebbing T., 2004. The effect of forage quality on N intake and N excretion under rotational grazing. In: Proceedings of the European Grassland Society, 21-24 June, Luzern, Switzerland, 675-677.
Schellberg J., Sudekum K.H. & Gebbing T., 2007. Effect of herbage on N intake and N excretion of suckler cows. Agron. Sustain. Dev., 27, 3030 -311.
Steinshamn H. et al., 2006. Feed nitrogen conversion in lactating dairy cows on pasture as affected by concentrate supplementation. Anim. Feed Sci. Technol., 131, 25-41.
Stevens R.J. & Laughlin R.J., 1997. The impact of cattle slurries and their management on ammonia and nitrous oxide emissions from grassland. In: Jarvis S.C. & Pain B.P., eds. Gaseous nitrogen emissions from grasslands. Wallingford, UK: CAB International, 233-256.
Van Keulen J. & Young B.A., 1977. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci., 44, 282-287.
Pour citer cet article
