
Bulk segregant analysis and marker-trait association reveal common AFLP markers for resistance to septoria leaf blotch in Tunisian old durum wheat
Received on April 29, 2013; accepted on February 3, 2014
Résumé
Les méthodes d’analyse de ségrégation en mélange et d’association marqueur-trait révèlent des marqueurs AFLP communs de résistance à la septoriose chez un ancien blé dur de Tunisie. La septoriose, causée par le champignon Mycosphaerella graminicola, est actuellement la maladie foliaire du blé dur la plus répandue en Tunisie. La plupart des cultivars élites tunisiens montrent peu ou pas de résistance à ce pathogène. Récemment, certaines variétés tunisiennes dont ‘Agili’ ont été identifiées comme hautement résistantes au pathotype virulent, ‘Tun06’. Afin d'identifier les marqueurs AFLP de résistance à la septoriose, nous avons utilisé l'analyse de ségrégation en mélange dans deux populations biparentales F3 issues de croisements entre l’accession résistante de blé dur ‘Agili’ et les cultivars sensibles ‘Karim’ et ‘Khiar’, et la cartographie d’association en utilisant une collection de 110 accessions anciennes de blé dur. Sur les 24 combinaisons d'amorces EcoRI/MseI testées, 4 ont été polymorphes entre les pools d'ADN résistants et sensibles. L’analyse du polymorphisme AFLP utilisant ces combinaisons d'amorces sur les parents et les 9 à 12 descendances de chaque mélange a révélé que les allèles, EcoACT/MseCTC-241, EcoACA/MseCTT-128 et EcoAGG/MseCTC-104 sont spécifiques aux individus résistants suggérant leur liaison avec le gène de résistance à la septoriose. Une régression logistique binaire sur la population locale de blé dur a montré que deux des marqueurs AFLP identifiés par BSA ont été significativement associés à la résistance. L'allèle EcoACT/MseCTC-241 a été significativement associée (P < 7.10-8) et située à une distance génétique de 1.2 cM du gène de résistance. Ce résultat indique l'efficacité de l'utilisation des populations locales pour détecter les marqueurs potentiels de résistance de nature qualitative.
Abstract
Septoria tritici blotch (STB) caused by the fungus Mycosphaerella graminicola is currently the major foliar disease of durum wheat in Tunisia. Most of the Tunisian elite cultivars show little or no resistance to this pathogen. Recently, some Tunisian landraces including ‘Agili’ have been identified as highly resistant to the virulent STB pathotype, ‘Tun06’. In order to identify amplified fragment length polymorphism (AFLP) markers associated with resistance to STB, we used bulk segregant analysis (BSA) in two F3 biparental populations derived from crosses between the resistant durum wheat accession ‘Agili’ and the susceptible cultivars ‘Karim’ and ‘Khiar’ and Association Mapping (AM) in a collection of 110 old durum wheat landraces. Out of the 24 EcoRI/MseI primer combinations tested, 4 were polymorphic between resistant and susceptible DNA pools. Screened on the parents and 9 to 12 progenies from each bulk, these polymorphic AFLP primer combinations revealed EcoACT/MseCTC-241, EcoACA/MseCTT-128, and EcoAGG/MseCTC-104 as specific alleles to resistant individuals suggesting their linkage to a gene for resistance to STB. Binary logistic regression analysis on the diversity panel showed that two AFLP markers out of the three identified by BSA were significantly associated with the resistance. EcoACT/MseCTC-241 was the significantly associated allele at P < 7.10-8 and was identified at a genetic distance of 1.2 cM to the resistance gene. This result indicates the efficiency of using landrace populations to detect potential markers for qualitative resistance.
1. Introduction
1Septoria tritici blotch (STB) caused by the ascomycete Mycosphaerella graminicola (anamorph Zymoseptoria tritici) is the primary fungal disease that affects durum wheat throughout the area around the Mediterranean Sea (Boukef et al., 2012). Chemical control is currently one of the main approaches used to manage STB (Jorgensen et al., 2008). However, fungicide resistance and its associated environmental impact is now a widespread problem, thereby necessitating genetic wheat improvement and the development of resistant varieties of wheat (Jorgensen, 2008; Cools et al., 2012). Selection for STB resistance can be facilitated by tagging effective genes with molecular markers that can subsequently be used in breeding programs to trace the useful alleles (Varshney et al., 2007). These markers have traditionally been identified through segregation analyses using bi-parental populations. Bulked segregant analysis (BSA), which consists of comparing two pooled DNA samples of individuals from a segregated population originating from a single cross, is considered to be a quicker method for identifying potential markers linked to monogenic qualitative trait (Michelmore et al., 1991). Recently, a new approach has been suggested that involves genotype-phenotype correlations in multiparental populations with broader genetic diversity (Flint-Garcia et al., 2005; Zhu et al., 2008; Hall et al., 2010). This approach, known as association mapping (AM) or linkage disequilibrium (LD) mapping, successfully addresses the challenge of detection and exploitation of DNA polymorphisms in plant systems (Ingvarsson et al., 2011). The use of multiparental populations presents clear advantages over biparental populations, and the two are often applied in conjunction to further validate the associations and to reduce the spurious ones (Tommasini et al., 2007).
2An important prerequisite for AM is the availability of accessions with suitable levels of genetic variation for target traits (Maccaferri et al., 2006). Several marker-trait association analyses have shown the greater efficiency of wild relative/landrace populations compared to modern cultivar panels, especially for mapping disease resistance genes (Jing et al., 2007; Mazzucato et al., 2008). In fact, genetic diversity of modern wheat germplasm is shown to be significantly lower than that of landrace accessions (Roussel et al., 2004; Hao et al., 2006). Therefore, wheat research targeting disease resistance improvement considers wild relative/landrace populations to be highly valuable resources for introducing new resistance genes and for better marker-trait detections and mapping resolution (Sodkiewicz et al., 2004). Several markers linked to STB resistance genes have been identified in bread wheat using BSA, including restriction fragment length polymorphism (RFLP), simple repetitive sequence (SSR) and amplified fragment length polymorphism (AFLP) (Adhikari et al., 2003; McCartney et al., 2003; Adhikari et al., 2004a; Adhikari et al., 2004b). To date, 18 major stb genes and QTLs for resistance to STB in bread wheat have been mapped (Tabib-Ghaffary, 2011). In durum wheat, resistance to STB has never been investigated at the genetic level until the recent identification of a single dominant gene conferring total resistance in the adult stage in ‘Agili’, an old Tunisian durum wheat accession (Ferjaoui et al., 2011). In addition, Gharbi et al. (2011) identified accessions tolerant to STB in Tunisian elite durum wheat germplasm.
3The aim of the present study was to identify potential AFLP markers for resistance to STB disease in durum wheat in the adult stage using two approaches:
4– BSA in segregating F3 progenies generated from crosses between a resistant accession ‘Agili’ and two susceptible cultivars ‘Khiar’ and ‘Karim’;
5– AM in a diverse old local durum wheat collection harboring phenotypic variation for traits including resistance to STB.
6The reliability of marker-trait association analysis would be the identification of the same markers linked to the gene of interest. Investigation of landrace populations to detect major genes for resistance is discussed.
2. Materials and methods
2.1. Plant material
7F2-derived F3 families, originating from two crosses between a resistant durum wheat parent ‘Agili’ and two susceptible durum wheat modern varieties ‘Khiar’ and ‘Karim’, were used for the BSA approach. These biparental populations consisted of 8 to 12 homozygous resistant and 9 to 13 homozygous susceptible progenies. In the AM analysis, the diversity panel used consisted of 9 completely resistant and 101 susceptible accessions of old durum wheat landraces. The resistance to STB was evaluated in the adult stage on both populations. Inoculation and disease evaluation methods are described below.
2.2. Inoculum preparation and plant inoculation
8‘Tun6’, the most virulent pathotype of STB in Tunisia, was used in this study (Medini et al., 2008). Tested on the Canadian differential hosts, this pathotype has been characterized by its virulence on three differential durum wheat lines of four tested. Inoculum was prepared by inoculating 250 ml of liquid yeast-glucose media (10 g of yeast extract and 30 g of glucose in 1 l of distilled water) in 500 ml Erlenmeyer flasks with fresh Z. tritici colonies in solid yeast glucose media containing agar (20 g·l-1). The Erlenmeyer flasks were incubated for 7 to 10 days with shaking (100 rpm). The resulting inoculum suspensions were filtered and adjusted to 107 spores per ml with distilled water. Ten drops of Tween 20 (polyoxyethylene-sorbitanmonolaurate) were added per liter of spore suspension to reduce surface tension. Plants were inoculated twice in the field at stem elongation (Zadoks scale 37) with a motor sprayer.
2.3. Disease assessment
9Symptoms of STB were assessed 28 days after the second inoculation (dpi). Susceptibility and resistance were measured using a qualitative scale, i.e., plants were scored as susceptible if leaves were covered by necrotic lesions bearing pycnidia, or as resistant if leaves of the whole plant had no pycnidia. F3 families were classified as homozygous resistant when all plants within the family were resistant, heterozygous when the family segregates for resistance, and homozygous-susceptible when all plants within the family were susceptible (Ferjaoui et al., 2011).
2.4. DNA extraction
10For both BSA and AM populations, total genomic DNA was isolated from leaf tissues of adult plants using a modified CTAB (hexadecyltrimethyl ammonium bromine) method (Saghai-Maroof et al., 1994). DNA extracted was visualized on 1% agarose gel for quality assessment, quantified on a spectrophotometer at a wavelength of 260 nm and adjusted to a final concentration of 50 ng·μl-1.
2.5. Bulk segregant analysis
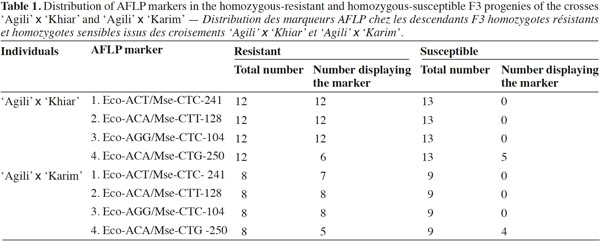
11Bulked segregant analysis (BSA) was performed as described by Michelmore et al. (1991). Nine to 12 homozygous-resistant and 9 to 12 homozygous-susceptible individuals of each F3 progeny were randomly selected to construct resistant and susceptible bulks (Table 1), respectively. Each bulk was at a final concentration of 50 ng·μl-1. Band patterns of bulks were compared with those from the parents to identify potential markers linked to the resistance.
2.6. AFLP procedure
12The AFLP procedure was conducted according to the protocol developed by Vos et al. (1995). Genomic DNA was digested with EcoRI and MseI at 37 °C for 4 h. Digested DNA fragments were ligated to EcoRI and MseI adapters with T4 DNA ligase at 16 °C for 12 h. After ligation, the reaction mixture was used as template DNA for pre-amplification. Pre-amplification was performed using primers with one selective nucleotide, followed by selective amplification using primers with three selective nucleotides. The reactions were conducted in a 25 µl reaction containing 2.5 ml 10× PCR buffer, 0.2 mMdNTPs, 1U Taq polymerase, and 0.5 µM EcoRI fluorescent-labeled selective primer (Applied Biosystems) and 1 µM MseI selective primer. Selective amplification was initiated by a denaturation step at 94 °C for 2 min, followed by 10 cycles of denaturation at 94 °C for 20 sec, annealing at 65 °C for 30 sec and extension at 72 °C for 2 min, and then followed by 20 cycles of denaturation at 94 °C for 30 sec, annealing at 56 °C for 30 sec, and extension at 72 °C for 2 min, and a final extension at 72 °C for 10 min. PCR products were analyzed using capillary electrophoresis ABI PRISM 3130 (Applied Biosystems).
2.7. Linkage, marker trait association, and linkage disequilibrium
13The potential markers identified by BSA were scored in each homozygous resistant and susceptible individual of each F3 progeny. Mapping was performed considering the population as RILs because the homozygous progenies were fixed for the resistance locus. Linkage analysis was performed with the QTL IciMapping version 3.1 (www.isbreeding.net). The Kosambi mapping function was used to convert recombination data into centimorgans (cM).
14The association between molecular markers and resistance to STB was determined by the procedure of binary logistic regression based on binary disease score and the AFLP markers using the SPSS software package (version 9.1). The AFLP allele (1; 0) was treated as an indicator variable and the phenotype as dependent variable. We fitted a multiple regression model of the trait response on the set of the AFLP allele treated as the indicator. To control for multiple testing, the correction of Bonferroni (http://mathworld.wolfram.com/BonferroniCorrection.html) was applied for the markers that were associated at P < 0.05. Markers with probabilities of associations less than the Bonferroni threshold of 0.00035 were retained.
15Linkage disequilibrium was estimated between pairs of AFLP markers that were significantly associated with the resistance using r2 coefficient (Pritchard et al., 2001), with the GDA software, version 1.1 (Lewis et al., 2001). Significant association was determined using a chi-square test. Pairs of markers that have P < 0.001 were declared significantly associated.
2.8. Segregation analysis of the resistance
16In order to assess the independence of STB resistance genes, crosses between the resistant accession ‘Agili’ and seven other resistant landraces identified were carried out. Inoculations using ‘Tun06’ pathotype were performed according to the methodology described above. Accordance of the observed segregations to 1(resistant):0(susceptible) expectation was tested using a chi-square test to detect possible distortion. A 1:0 ratio is expected when identical resistance genes are involved; a 15:1 ratio is expected for two clearly distinct dominant resistance genes in the parent.
3. Results
3.1. Bulk Segregant Analysis (BSA)
17Twenty-four primer combinations were assessed on homozygous-resistant and homozygous-susceptible pools of the F3 families derived from the crosses ‘Agili’ (resistant) x ‘Khiar’ (susceptible) and ‘Agili’ x ‘Karim’ (susceptible). The total number of bands varied from 27 (for EcoAGC/MseCAG primer combination) to 155 (for EcoACA/MseCTC primer combination). The comparison between resistant and susceptible bulks of each cross revealed that four primer combinations produced a distinct peak (band) in resistant bulks and in resistant parents and was absent in susceptible bulks and susceptible parents, indicating their linkage with resistance trait. These four primer combinations were further analyzed for segregation in individuals of resistant and susceptible bulks. Only three primer combinations produced a peak (marker band) specific to the resistant bulk (namely, EcoACT/MseCTC-241, EcoACA/MseCTT-128, and EcoAGG/MseCTC-104), co-segregated with the resistance in ‘Agili’ x ‘Khiar’ progenies, whereas the marker EcoACA/MseCTG-250 did not. The allele EcoACT/MseCTC-241 was absent in one resistant progeny of ‘Agili’ x ‘Karim’, allowing us to estimate recombination distance between the resistance locus and the AFLP marker. Linkage analysis of the F3 progenies showed that EcoACT/MseCTC-241 mapped at 1.2 cM from the stb gene.
3.2. Marker-trait association and linkage disequilibrium
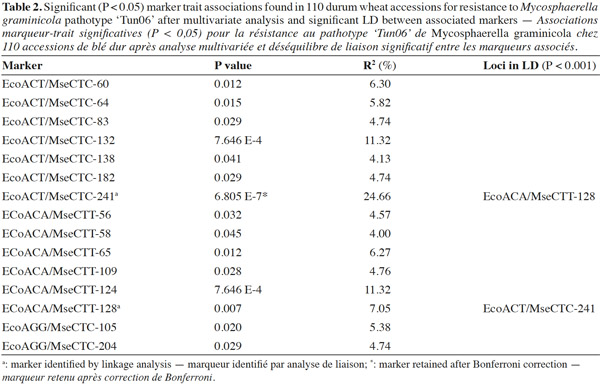
18The three successful primer combinations that identified three potential markers for resistance to leaf blotch by BSA were assessed on a collection of 9 fully resistant and 101 susceptible accessions to the M. graminicola isolate ‘Tun06’. Multivariate analysis showed that 15 markers from a total of 110 were associated at P < 0.05 with the resistance (Table 2). The marker EcoCT/MseCTC-241 was associated at a high significant probability (P < 7·10-8). This marker was retained after Bonferroni correction. The EcoACA/MseCTT-128 marker was associated with resistance but at a lower significance level than the marker EcoACT/MseCTC-241. In fact, the EcoACA/MseCTT-128 allele was present in three susceptible accessions, whereas the EcoACT/MseCTC-241 allele was present in only one susceptible accession (Table 3). The presence of the resistance marker allele (band) in the susceptible progenies could be originated from another part of the genome and co-migrated with identical mobility to the resistant marker allele. In addition, the EcoACA/MseCTT-128 allele was found in significant linkage disequilibrium (LD) with EcoACT/MseCTC-241 as demonstrated above by linkage analysis (Table 2). However, marker trait association analysis did not show an association between resistance and EcoAGG/MseCTC-104 allele as revealed by linkage analysis. In fact, the marker EcoAGG/MseCTC-104 was found in 42 susceptible accessions to the isolate ‘Tun06’.

4. Discussion
19For the past 20 years, investigation for disease resistance has been an important objective in molecular breeding of wheat. In this study, two approaches were used to identify AFLP markers linked to a STB resistance gene:
20– BSA in biparental F3 populations derived from crosses between a resistant accession ‘Agili’ and two susceptible cultivars, ‘Khiar’ and ‘Karim’;
21– association mapping in durum wheat landraces with a highly diverse genetic background.
22Both strategies identified two reliable AFLP markers linked to the stb gene that provides a qualitative resistance against the most frequent pathotype of M. graminicola reported in Tunisia.
23Molecular markers linked to disease resistance traits have great importance but are difficult to reveal especially when the disease is difficult to score due to the multi-component of pathogen fitness and when the trait has a small effect on phenotypic variation and is influenced by environmental factors (Sukhwinder-Singh et al., 2012). In addition, available genetic resources are difficult to obtain and are not always efficient in detecting loci underlying qualitative/quantitative traits. This study provides evidence that the existing association panel and disease assessment procedure can be used to detect AFLP markers for STB resistance once the trait is qualitative. Despite the small population size, low frequency of the resistant phenotypes in the tested population (less than 10%), and not taking into consideration population structure, the association study was conclusive and in agreement with the BSA results. This is attributed to the qualitative approach of the resistance and its simple interpretation using a binary disease scoring (1: susceptibility; 0: resistance) that enable the use of binary logistic regression to analyze the association. Here, common issues related to marker-trait association analysis are avoided, e.g., false positive associations caused by population subdivision (Pritchard et al., 2001) and limited power to detect rare loci within populations (Flint-Garcia et al., 2005; Breseghello et al., 2006; Soto-Cerda et al., 2012).
24The AFLP markers identified by association analysis were at a minimal distance of 1.2 cM from stb gene. This genetic distance is not significant due to small number of segregating individuals and may increase by enhancing the population size. However, we can assume that a minimal interval of 2.4 cM (LD extent) is required between markers for association mapping using this durum wheat landrace population. This low LD may probably inhibit marker-aided selection and thus require a large number of markers to detect an associated region. The level of LD depends on population structure, cross/self pollination ratios, effective population size, and selection pressure and may vary across genome regions (Maccaferri et al., 2005). The LD extent observed here is similar to the LD pattern described in landrace accessions compared to wheat-inbred lines (Caldwell et al., 2006). The extent of LD less than 1 cM was described in landrace populations because of high recombination events in the ancestry of the accessions (Comadran et al., 2009). However, the average LD within 189 cultivars and advanced lines of durum wheat accession from the Mediterranean area was decayed over a distance of 10 cM due to gene selection during plant breeding programs (Maccaferri et al., 2006),
25In this study, seven resistant accessions lacked the identified AFLP markers for the resistance to STB. Two hypothetical explanations can be suggested:
26– the linkage could have been broken between the marker and the stb gene within these resistant accessions due to recombination events after the stb gene introgression;
27– these resistant accessions may carry different resistance alleles from the ones identified in ‘Agili’.
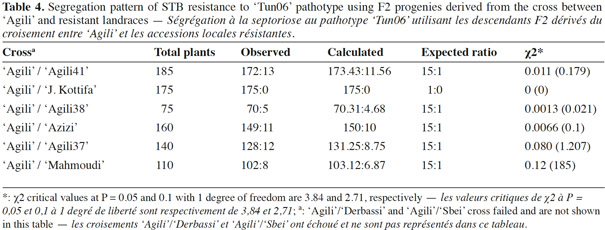
28The allelic test for resistance to the ‘Tun06’ pathotype using F2 progenies derived from the cross between ‘Agili’, and these resistant accessions revealed susceptible individuals. The segregation pattern of STB resistance was approaching a 15:1 ratio in the cross, confirming the presence of two clearly distinct dominant resistance genes in the parent (Table 4). The ‘Agili’ x ‘JenahKhotifa’ cross was the exception, where no susceptible progeny was shown indicating the presence of identical resistance genes in the parent. ‘Agili’ and ‘JenahKhotifa’ landrace accessions, which share the EcoACT/MseCTC-241, EcoACA/MseCTT-128 and EcoAGG/MseCTC-104 alleles, probably represent common ancestry. The resistance genes identified likely derive from wild relatives and were introgressed through natural outcrossing events into the durum wheat landraces. Evidence of natural gene flow between wheat and wild relatives have been detected (Arrigo et al., 2011), as well as several disease resistance genes to rusts, powdery mildew and fusarium head blight in domesticated wheat genome cultivars derived from wild relatives including Aegilops and Triticum species and other diploid wheat grasses (Monneveux et al., 2000; Jauhar et al., 2009). These findings emphasize the need for further study of resistance mechanisms in order to fully exploit the landrace reservoir for STB resistance in cultivated durum wheat.
5. Conclusion
29Finally, this study demonstrates relevant preliminary results in identifying STB resistance genes and leads us to increase the research interest in pursuing advanced studies. The next step will be to move towards more precise localization of the identified stb resistance gene by using a large population of recombinant inbred lines in order to successfully apply marker-assisted selection in breeding program for durum wheat. In addition durability of the resistance and the evolutionary potential of the pathogen, major components for a successful virulence management, should be considered. Potentially durable resistance genes are valuable tools for breeding, especially if combined with other stb genes and/or resistance QTLs as part of a pyramiding strategy.
30Acknowledgements
31This study was supported by Tunisian government funded project untitled “Breeding for resistance to biotic stress in durum wheat”. We thank the National Gene Bank for providing facilities for conducting AFLP genotyping.
Bibliographie
Adhikari T.B., Anderson J.M. & Goodwin S.B., 2003. Identification and molecular mapping of a gene in wheat conferring resistance to Mycosphaerella graminicola. Phytopathology, 93(9), 1158-1164.
Adhikari T.B., Wallwork H. & Goodwin S.B., 2004a. Microsatellite markers linked to the Stb2 and Stb3 genes for resistance to septoria tritici blotch in wheat. Crop Sci., 44(4), 1403-1411.
Adhikari T.B. et al., 2004b. Molecular mapping of Stb1, a potentially durable gene for resistance to septoria tritici blotch in wheat. Theor. Appl. Genet., 109(5), 944-953.
Arrigo N. et al., 2011. Gene flow between wheat and wild relatives: empirical evidence from Aegilops geniculata, Ae. neglecta and Ae. triuncialis. Evol. Appl., 4(5), 685-695.
Boukef S. et al., 2012. Frequency of mutations associated with fungicide resistance and population structure of Mycosphaerella graminicola in Tunisia. Eur. J. Plant Pathol., 132(1), 111-122.
Breseghello F. & Sorrells M.E., 2006. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics, 172(2), 1165-1177.
Caldwell K.S., Russell J., Langridge P. & Powell W., 2006. Extreme population-dependent linkage disequilibrium detected in an inbreeding plant species, Hordeum vulgare. Genetics, 172(1), 557-567.
Comadran J. et al., 2009. Patterns of genetic diversity and linkage disequilibrium in a highly structured Hordeum vulgare association-mapping population for the Mediterranean basin. Theor. Appl. Genet., 119(1), 175-187.
Cools H.J. & Fraaije B.A., 2012. Update on mechanisms of azole resistance in Mycosphaerella graminicola and implications for future control. Pest Manage. Sci., 69(2), 150-155.
Ferjaoui S., Sbei A., Aouadi N. & Hamza S., 2011. Monogenic inheritance of resistance to septoria tritici blotch in durum wheat ‘Agili’. Int. J. Plant Breed., 5, 17-20.
Flint-Garcia S.A. et al., 2005. Maize association population: a high-resolution platform for quantitative trait locus dissection. Plant J., 44(6), 1054-1064.
Gharbi M.S., Berraies S., Ammar K. & Yahyaoui A., 2011. Delivering disease resistance to enhance sustainability of durum wheat production in Tunisia. In: Duveiller E. & Pawan K.S., eds. Proceedings of the 8th International Symposium on Mycosphaerella and Stagonospora diseases of cereals, 11-14 September, 2011, Mexico City, Mexico. Mexico City, Mexico: CIMMYT, 60.
Hall D., Tegstrom C. & Ingvarsson P.K., 2010. Using association mapping to dissect the genetic basis of complex traits in plants. Briefings Funct. Genomics, 9(2), 157-165.
Hao C.Y. et al., 2006. Genetic diversity and core collection evaluations in common wheat germplasm from the northwestern spring wheat region in China. Mol. Breed., 17(1), 69-77.
Ingvarsson P.K. & Street N.R., 2011. Association genetics of complex traits in plants. New Phytol., 189(4), 909-922.
Jauhar P.P., Peterson T.S. & Xu S.S., 2009. Cytogenetic and molecular characterization of a durum alien disomic addition line with enhanced tolerance to Fusarium head blight. Genome, 52(5), 467-483.
Jing H.-C. et al., 2007. Identification of variation in adaptively important traits and genome-wide analysis of trait-marker associations in Triticum monococcum. J. Exp. Bot., 58(13), 3749-3764.
Jorgensen L.N., 2008. Resistance situation with fungicides in cereals. Zemdirbyste-Agriculture, 95, 373-378.
Jorgensen L.N. et al., 2008. Integrating disease control in winter wheat optimizing fungicide input. Outlook Pest Manage., 19, 206-213.
Lewis P.O. & Zaykin D., 2001. Genetic data analysis. Computer program for the analysis of allelic data, Version 1.0 (d16c), http://www.eeb.uconn.edu/people/plewis/software.php, (18.02.14).
Maccaferri M., Sanguineti M.C., Noli E. & Tuberosa R., 2005. Population structure and long-range linkage disequilibrium in a durum wheat elite collection. Mol. Breed., 15(3), 271-290.
Maccaferri M. et al., 2006. A panel of elite accessions of durum wheat (Triticum durum Desf.) suitable for association mapping studies. Plant Genet. Resour., 4, 79-85.
Mazzucato A. et al., 2008. Genetic diversity, structure and marker-trait associations in a collection of Italian tomato (Solanum lycopersicum L.) landraces. Theor. Appl. Genet., 116(5), 657-669.
McCartney C.A., Brulé-Babel A.L., Lamari L. & Somers D.J., 2003. Chromosomal location of a race-specific resistance gene to Mycosphaerella graminicola in the spring wheat ST6. Theor. Appl. Genet., 107(7), 1181-1186.
Medini M. & Hamza S., 2008. Pathotype and molecular characterization of Mycosphaerella graminicola isolates collected from Tunisia, Algeria and Canada. J. Plant Pathol., 90(1), 65-73.
Michelmore R.W., Paran I. & Kesseli R.V., 1991. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. U.S.A., 88(21), 9828-9832.
Monneveux P., Zaharieva M. & Rekika D., 2000. The utilisation of Triticum and Aegilops species for the improvement of durum wheat. In: Royo C., Nachit M., Di Fonzo N. & Araus J.L., eds. Durum wheat improvement in the Mediterranean region: new challenges. Zaragoza, Spain: CIHEAM, Options Méditerranéennes: Série A. Séminaires Méditerranéens; n° 40, 71-81.
Pritchard J.K. & Przeworski M., 2001. Linkage disequilibrium in humans: models and data. Am. J. Hum. Genet., 69(1), 1-14.
Roussel V., Koenig J., Beckert M. & Balfourier F., 2004. Molecular diversity in French bread wheat accessions related to temporal trends and breeding programmes. Theor. Appl. Genet., 108, 920-930.
Saghai Maroof M.A. et al., 1994. Extraordinarily polymorphic microsatellite DNA in barley: species diversity, chromosomal locations, and population dynamics. Proc. Natl. Acad. Sci. U.S.A., 91(12), 5466-5470.
Sodkiewicz W. & Strzembicka A., 2004. Application of Triticum monococcum for the improvement of triticale resistance to leaf rust (Puccinia triticina). Plant Breed., 123(1), 39-42.
Soto-Cerda B.J. & Cloutier S., 2012. Association mapping in plant genomes. In: Çalişkan M., ed. Genetic diversity in plants. Rijeka, Croatia: InTech, 29-53.
Sukhwinder-Singh et al., 2012. Multi-trait and multi-environment QTL analyses for resistance to wheat diseases. PLoS ONE, 7(6), e38008, doi:10.1371/journal.pone.0038008.
Tabib Ghaffary S.M., 2011. Efficacy and mapping of resistance to Mycosphaerella graminicola in wheat. PhD thesis: University of Wageningen (The Netherlands).
Tommasini L. et al., 2007. Association mapping of Stagonospora nodorum blotch resistance in modern European winter wheat varieties. Theor. Appl. Genet., 115(5), 697-708.
Varshney R. et al., 2007. Genic molecular markers in plants: development and applications. In: Varshney R. & Tuberosa R., eds. Genomics-assisted crop improvement. Vol. 1. Genomics approaches and platforms. Dordrecht, The Netherlands: Springer, 13-29.
Vos P. et al., 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res., 23(21), 4407-4414.
Zhu C., Gore M., Buckler E.S. & Yu J., 2008. Status and prospects of association mapping in plants. Plant Genome, 1(1), 5-20.
Pour citer cet article
